|
Blood Group Antigens and Cell Adhesion
|
|
|
 |
Blood group antigens are the molecules expressed on the surface of human red blood cells (RBC), against which antibodies occur naturally or can be raised. The most popular blood group antigens of clinical importance are the blood-group-ABO antigens and Rh antigens. Blood group antigens comprise a large number of molecular epitopes including polypeptides, glycoconjugates and glycoproteins (1). The most famous blood group glycoantigens and their related glycoantigens are listed in Fig. 1. The human blood group A or B antigens are present on RBC as glycolipids and as glycoproteins. The A-antigen and the B-antigen are trisaccharides, and the H antigen is a disaccharide, which is the backbone of A/B trisaccharides. The ABO-blood-group antigens are found on glandular epithelial cells and other epithelial cells including endothelial cells. They were found also on von Willebrand factor the blood coagulation system. The presence of the antigens on this factor may cause problems in treating hemophilia. In a secretor, the antigens are also detected in saliva and other secretions. The ABO blood system is the classic example of multiple allelism. To the galactose of H-antigen, a galactose molecule is linked through alpha 1,3 linkage to form the blood-group-B trisaccharide. If N-acetylgalactosamine is transferred to the galactose of H-antigen , blood group A trisaccharide is formed. The genetic locus for the glycosyltransferase for synthesizing the trisaccharides is located on chromosome 9 in human, and the difference in the primary structure of the transferase changes the specificity of the enzyme. For example, specific differences in four amino acids in the glycosyltransferase coded in the region give the protein the activity of galactosyltransferase, while the other combination of amino acids in the four amino acids changes the protein into N-acetylgalactosaminyltransferase. When the enzyme loses the glycosyltransferase activity by nonsense mutation in the genetic locus, the enzyme cannot transfer glycoepitopes at all and the result is that only the blood-group-H antigens are expressed on RBC.
Lewis a and Lewis b antigens are blood group antigens related to blood-group-ABO antigens (Fig.1). Lewis a antigen was first recognized by an antibody in a serum sample from Mrs. H.G.D.Lewis. The abnormal expression of the sialylated form of the Lewis a antigen (Sialyl Lea (Lewis a) ) is closely correlated with various forms of cancer including pancreatic cancer, gallbladder/bile duct cancer and cholangiocarcinoma. A monoclonal antibody CA19-9 against sialyl Lea is a popular diagnostic agent for these tumors. Note that this antibody is useless for cancer diagnosis when a patient is lacking the enzyme for the synthesis of sialyl Lea. In Japanese, about 5-10% of the population lacks this enzyme. Lex (Lewis x) antigen and its sialylated form (sialyl Lex antigen) are not blood-group-antigens but their structure is related to Lewis blood group antigens (Fig.1). The stage-specific embryonic antigens (SSEA-1) famous in developmental biology is the Lex antigen. The sialyl Lex is known as a receptor of E-selectin and is important in lymphocyte homing. In mouse embryos, before the 8-cell stage, cell-cell adhesion between blastomeres is not so tight that cell-cell boundaries are clearly visible. Late in the 8-cell stage, a process known as "compaction" begins, and cell-cell adhesion becomes very tight and cell-cell boundaries become indistinguishable. The compaction is blocked by the addition of Lex glycoconjugates, indicating that Lex plays an active role in calcium dependent cell-cell adhesion of mouse embryos. |
|
|
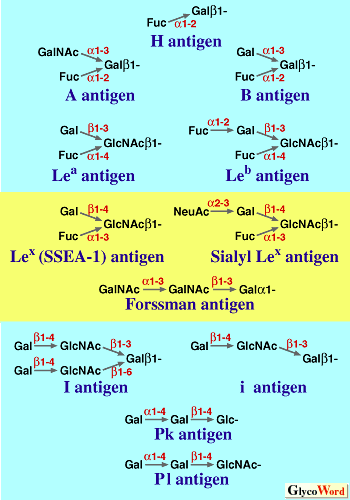
Major carbohydrate
blood group antigens
(pale blue)
and related antigens
(yellow) |
|
|
|
Cell adhesion molecules are divided into two different groups: cell-cell adhesion molecules and cell-substratum adhesion molecules. The latter are molecules mediating adhesion of cells onto extracellular matrices present outside cells. Of course, this grouping of cell adhesion molecules cannot be so seriously taken, and the possibility that one specific molecule (such as integrin) is acting as a cell-cell adhesion molecule and a cell-substratum adhesion molecule simultaneously must always be kept in mind. Cell-cell adhesion molecules are divided into two groups: calcium-dependent cell-cell adhesion molecules and calcium-independent cell-cell adhesion molecules. The most famous calcium-dependent cell-cell adhesion molecules are cadherin adhesion molecules. NCAMs are the best known calcium-independent cell-cell adhesion molecules. Studies on cell-cell adhesion molecules have been greatly facilitated through the introduction of monoclonal antibodies which disrupt cell-cell adhesion. All the major classical cadherins were identified by raising specific antibodies against various tissue cell lines which disrupt calcium-dependent cell-cell adhesion of the cells. For instance, E-cadherin was identified as a protein recognized by the monoclonal antibody ECCD-1 which disrupts calcium dependent cell-cell adhesion of mouse embryonic carcinoma cell line F9. N-cadherin and P-cadherin were also identified as epitopes recognized by the blocking antibodies of calcium-dependent cell-cell adhesion, NCD-1 and PCCD-1, respectively. The former antibody blocks calcium dependent adhesion of chicken neural retinal cells and the latter blocks adhesion of P9, which is the cell line-derived placenta of mouse cells. The antibody against E-cadherin (ECCD-1) reversibly blocks compaction of mouse embryos, suggesting that E-cadherin plays a role in mouse compaction. Addition of polyvalent sialyl Le x or anti-beta galactosyltransferase antibody also blocks compaction. This strongly indicates that not only cadherin but also glycoantigens play important roles in calcium-dependent cell-cell adhesion.
Since the use of blocking monoclonal antibodies in identifying cell-cell adhesion molecules was so effective, we also adopted this approach to identify calcium-dependent cell-cell adhesion molecules of frog (Xenopus laevis) blastula cells. The epitope recognized by one of the blocking antibodies was identified as human blood-group-B epitope. The blood group B epitopes were found on frog glycolipids in blastula membranes as well as on glycoproteins of blastula membranes. Calcium-dependent cell-cell adhesion of blastula cells was blocked by IgM type and IgG type anti-B monoclonal antibody, and by Fab of anti-B antibody. Anti-A and anti-H antibodies had no effect at all on cell adhesion. The adhesion was also blocked by blood-group-B antigen itself. Addition of blood group B-active mucopolysaccharides or blood group B active trisaccharides also disrupts calcium-dependent cell-cell adhesion but A-active and H-active substances were not effective. From these experiments, we concluded that human blood-group-B antigens play very important roles in calcium-dependent cell-cell adhesion of frog embryonic cells. The B-active glycolipids and glycoproteins were analyzed by our group and the molecular nature of the glycolipids and glycoproteins have been described (2). The B-active glycoproteins were found to be a lectin binding blood group B active substance. The possible interaction between the lectin and B active glycolipids in frog cell-cell adhesion is now being studied in our laboratory. In conclusion, in frog embryos, human-blood-group B antigens play a very important role as an indispensable member of cell-cell adhesion interaction. It is very important to know whether blood group ABO antigens also play similar roles in cell-cell adhesion of higher vertebrates. In this respect, a recent paper showing that the histo-blood type A glycosylation of integrin receptors is closely correlated with haptotactic motility and proliferation of tumor cells is intriguing (3). It is also very important to know whether evolutionary changes in glycosequences have affected their functions. |
|
|
|
Kazuko H. Nomura, Souhei Mizuguchi, Kazuya Nomura (Kyushu University, PRESTO, JST) |
|
|
|
|
|
| References |
(1) |
Reid, ME, Lomas-Francis, C, The Blood Group Antigen:Facts Book, Academic Press, New York, 1997 |
|
(2) |
Nomura, KH, et al. Dev. Genes Evol. 208, 9-18, 1998 |
|
(3) |
Ichikawa, D et al. Int. J. Cancer 76, 284-289, 1998 |
|
|
|
|
|
| Dec.15, 1998 |
|
|
|
|
|
|
|



