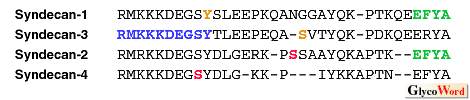|
|
Signal Transduction Mediated by Syndecans |
||||||||||||||||||||||||||||||||
 |
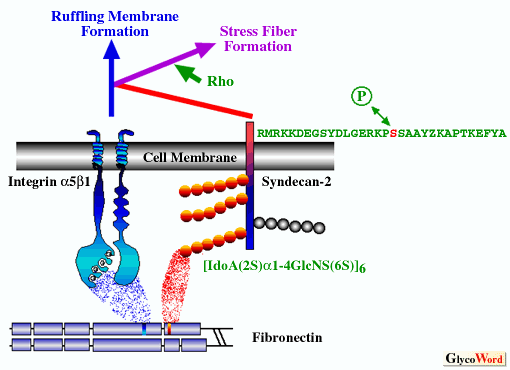
Recently much attention has focused on a signal transduction mediated by cell membrane-type proteoglycan, especially transmembrane heparan sulfate proteoglycan, syndecans. The present situation in this field is like a jigsaw puzzle in which some pieces are matched here and there, but the whole picture remains to be figured out.1,2) One of the difficulties in analyzing syndecans as signaling molecules may be attributed to their complex nature that a ligand-binding domain is a heparan sulfate side chains and a signaling moiety is thought to be a cytoplasmic portion of the protein core. What kind of signal is transduced through syndecans? The fact that the ligand-binding domain of syndecans is heparan sulfate side chains means that syndecans can act as receptors to any heparin-binding molecules. Actually, it has been shown that a variety of heparin-binding extracellular matrix components and growth factors bind to heparan sulfate side chains of syndecans. In most cases, there are other specific receptor proteins with higher binding affinity to these ligand molecules, therefore syndecans are thought to be low-affinity co-receptors that enhance or modify the signal transduction mediated by intrinsic receptors. Under these circumstances, the starting and ending points of the signal-cascades in which syndecans are involved are so ambiguous that it is hard to analyze accurate signaling of syndecans. Our study, which clearly shows that syndecan-2 acting as a co-receptor for a fibronectin substrate with integrin alpha5beta1 regulates signaling via integrin alpha5beta1 to alter cytoskeletal organizations, is a rare case in which the entrance and the exit of the cascade are defined (Figure). Data accumulated by a number of studies suggest that syndecans mediate the signal transduction for cytoskeletal organization required for cell adhesion and migration by acting as a receptor for extracellular matrix components and are required for cell growth by acting as a low-affinity receptor of growth factors. |
|||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| Redundancy and specificity of ligand-binding of syndecans Although the skeletal structure of heparan sulfate chains is composed of alternate saccharides of GlcA/IdoA and GlcNAc, it is calculated theoretically to be 1036 kinds of heparan sulfate chains by the number and positions of sulfate residues. In this context, it is hard to judge whether the explanation of "redundancy" in ligand-binding to heparan sulfate chains is a true redundancy or an apparent redundancy in which every binding is unique occurring through specific structures of heparan sulfate. The minimal domain structures of heparan sulfate chains required to bind different ligands, which have been determined a few cases, are slightly different from each other, indicating at least that heparan sulfate chains bind ligands via specific structures. If saccharide sequences having sulfate residues along heparan sulfate chains were determined like an amino acid sequence of a protein, this problem might be dissolved. However, at present the methods for analyzing posttranslational modifications have not been established, rendering the determination of ligand-binding specific structure of heparan sulfate very difficult. What protein is next to syndecans in the cascade? Amino acid sequences of the cytoplasmic domains of syndecan family members show a high degree of homology among the four members (Table). In particular, the membrane proximal part and C-terminus are highly conserved and the central part is unique to each member, suggesting that the former may be a trigger of a signal cascade common in the family and the latter may be the trigger of a distinctive cascade for each member. |
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| After phosphorylation of serine residue of the cytoplasmic domain of syndecan-2 was demonstrated in mouse lung carcinoma cells in situ,3) phosphorylation of other syndecans were sought. Phosphorylation of serine residue of syndecan-3 using a synthetic peptide, tyrosine residue of syndecan-1 using transfectants, and serine residue of syndecan-4 in fibroblast in situ were reported.4,5) However, the biological significance of phosphorylation/dephosphorylation in these amino acid residues has not been elucidated. Efforts to detect cytoplasmic proteins which interact with syndecans did not obtain any results for a long time. However, very recently it was demonstrated with a yeast two-hybrid screening method that the cytoplasmic portion of syndecans binds syntenin containing a tandem repeat of the PDZ domain 6) and that the PDZ domain of CASK binds syndecan-2.7,8) Syndecans bound to the PDZ domain with its C-terminus EFYA sequence, CASK was also bound to actin-binding protein 4.1 via it's another domain, and CASK colocalized to syndecan-1, suggesting that the binding ability to the PDZ domain is common to all family members and that the binding signal transducs to the cytoskeleton organization. It was also demonstrated that cortactin, an F-actin-binding protein which is a major substrate to Src kinase, bound to the immobilized cytoplasmic portion of syndecan-3.9) This binding was competed with the peptide of the membrane proximal common sequence of the syndecan cytoplasmic portion. At present there are fragmentary reports concerning syndecan signal transduction, but it is expected that these pieces of evidence will come together gradually and a whole picture of the puzzle will appear in the near feature. Nevertheless, it should be noted that nothing has been coprecipitated with syndecans by anti-syndecan antibodies in spite of every effort. This is attributed to the extraction conditions used. However, if this is a case of Sherlock Holmes' "the curious incident of the dog in the night-time," it will not be so easy to solve the problem. Dally, Drosophila homolog of glypican, another cell membrane-type heparan sulfate proteoglycan with a GPI anchor, has been detected in association with a mutation of abnormal cell division, and analyses of the signal cascade involved with glypican as a receptor of TGF-beta homolog, Dpp, are now actively being studied. Drosophila homolog of the syndecan gene has also been cloned. However, a mutant fly linking with syndecan has not been found out, thus an analysis has not progressed in this system either. If the mutant is found out, the information will contribute to understand signal transductions mediated by syndecans. |
||||||||||||||||||||||||||||||||
| Kayoko Oguri (Clinical Research Institute, National Nagoya Hospital) | ||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||
| Dec.15, 1998 | ||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||