|
|
|
|||||||||||||||||
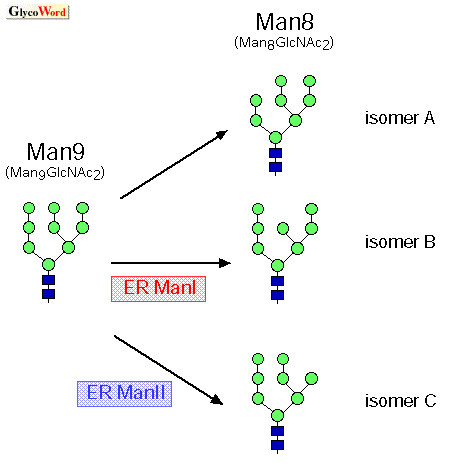
| Glycoprotein ERAD The roles of N-linked oligosaccharides on the ER quality control (ERQC) of newly synthesized glycoproteins are discussed in this series of GlycoWord, and in this session, I would like to introduce recently published data focusing on ER-associated degradation (ERAD) and mannose trimming from the N-linked glycans in the ER. Class I The mannose trimming from the N-linked oligosaccharides in the ER is shown in Fig. 1 as a very simplified schema. ER |
|||||||||||||||||
|
|||||||||||||||||
|
Mammalian EDEM (ER degradation enhancing
|
|||||||||||||||||
|
|||||||||||||||||
|
The roles of ER ManI and EDEMs in glycoprotein ERAD
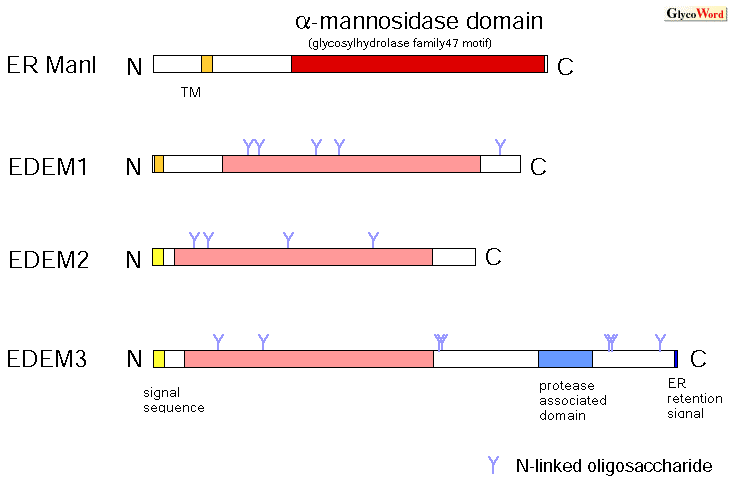
It was recently reported that each mammalian EDEMs (EDEM1, 2, 3) enhanced glycoprotein ERAD (1, 2, 3). ER ManI also accelerates the degradation of misfolded glycoproteins when they are transfected into mammalian cells. ER ManI is an enzyme which trims the mannose from the N-linked oligosaccharides in the ER, and this trimming becomes the signal for newly synthesized glycoproteins entering the degradation pathway from the productive folding cycle (refer to QS-A00, QS-A01). Although the schema of glycoprotein ERQC looks more valid by the addition of the molecules such as ER ManI and EDEMs, new questions arise and it is not so simple as the model shows. Functional analysis of EDEM3 gives rise to a new question as to why EDEM 1, 2, and 3 form a group distinct from ER ManI We have recently shown that EDEM3 enhances the ERAD of misfolded glycoproteins and mannose trimming from the N-linked oligosaccharides when it is transfected to mammalian cells. When we introduce a mutation to one of the conserved acidic amino acid residues of EDEM3, which is known to be important for the enzyme activity, mannose trimming from the glycoprotein is inhibited (3). We are now trying to detect the enzyme activity of EDEM3 in vitro. Because the mannosidase activity is not detected in yeast Htm1p/Mnl1p and in mammalian EDEM1, 2, these proteins are expected to work as lectins which recognize N-linked glycans, although the lectin activity has not yet been demonstrated in vitro. The next question then is why EDEMs including proteins with or without mannosidase activity form a group distinct from ER ManI, which apparently has enzyme activity. Our working hypothesis is that the |
|||||||||||||||||
| Nobuko Hosokawa (Institute for Frontier Medical Sciences, Kyoto University) | |||||||||||||||||
|
|||||||||||||||||
| Jun.5, 2006 | |||||||||||||||||
|
|
|||||||||||||||||
|
|||||||||||||||||