

|
|
Peptide:N-glycanase (PNGase) and free N-glycans in cytosol |
|||||||||||||||||||||||
|
Cytoplasmic Peptide:N-glycanase Peptide:N-glycanase (PNGase) releases N-glycans from glycoproteins/glycopeptides, and this enzyme from bacteria and plants has been extensively used as a powerful "tool" reagent to study the structure and biological functions of N-linked glycans on glycoproteins. The cytoplasmic PNGases, ubiquitously found throughout eukaryotes, are now widely recognized as a component implicated in the ERAD (ER-associated degradation) process, which constitute one of the quality control mechanisms for newly synthesized misfolded glycoproteins exported from the ER lumen. The enzyme is classified as a transglutaminase-superfamily that contains a putative catalytic triad of amino acids (cysteine, histidine, and aspartic acid). The mammalian orthologues of PNGase contain the N-terminal PUB domain, which serves as a protein-protein interaction domain, as well as the C-terminus domain, which has recently been found to be a novel carbohydate-binding domain. These structural features indicate the sophisticated coordination of this protein in complex formation, as well as substrate recognition. For more details on the structure and functions of this enzyme, the reader is directed to our recent reviews (1-3). |
|||||||||||||||||||||||
|
Free N-linked glycans: formation and degradation The occurrence of PNGase in cytosol and its participation in the ERAD process is obvious, but one question arises : what will be the fate of "free" N-glycans that are released by PNGase in the cytosol? Since the discovery of the ERAD process, extensive studies have focused on the fate of misfolded proteins. Curiously enough, however, not much attention has been paid with regard to the molecular mechanism and importance of free oligosaccharide metabolism in cytosol. In mammalian cells, so far two cytosolic glycosidases have been identified to be involved in the processing of free oligosaccharides: endo-β-N-acetylglucosaminidase (ENGase) and α-mannosidase (Man2C1) (Figure 1). It should be noted here that there are still many more enzymes/transporters that remain to be identified, although this "non-lysosomal" metabolic path for N-glycans may represent one of the very basic biological phenomena in eukaryotes (Figure 1). For further reading on this topic, the reader is directed to our recent review (4). |
|||||||||||||||||||||||
|
|||||||||||||||||||||||
| Tadashi Suzuki (Department of Biochemistry and 21st COE(Center of Excellence) Program, Osaka University Graduate School of Medicine; CREST (Core Research for Evolutionary Science and Technology), JST (Japan Science and Technology Agency)) |
|||||||||||||||||||||||
|
|||||||||||||||||||||||
| Apr. 19, 2007 | |||||||||||||||||||||||
|
|
|||||||||||||||||||||||
|
|||||||||||||||||||||||
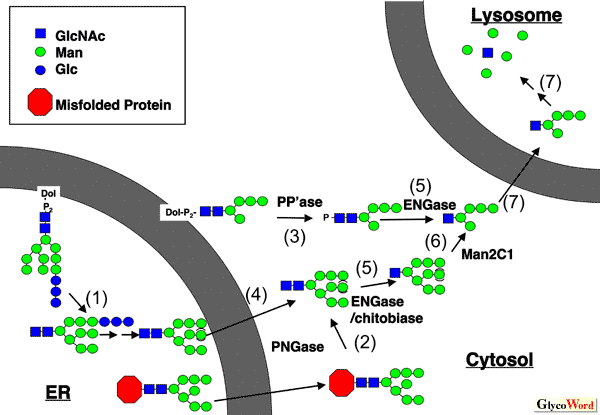
 -glucosidase I and II (and ER
-glucosidase I and II (and ER  -mannosidases. This non-lysosomal degradation pathway has been deduced mainly from the structures of free oligosaccharides that have been biochemically isolated.
-mannosidases. This non-lysosomal degradation pathway has been deduced mainly from the structures of free oligosaccharides that have been biochemically isolated.
