|
Sialic Acid and Evolution
|
|
|
 |
Introduction |
| Sialic acid has been defined as a neuraminic acid (5-amino-3,5-dideoxy D-glycero-D-galacto nonurosonic acid, Neu) and its derivatives 1). Since KDN whose aminoacyl group at the C5-position is substituted for the hydroxyl group was found in 19862), sialic acid was redefined as a nine-carbon sugar containing α-keto acid group (2-keto-3-deoxynononic acid). Sialic acid consists of acetylated, sulfated, methylated, and lactylated derivatives and is a large family of more than 50 members3). Diversity in sialic acid structure is a characteristic feature and such diversity is not observed in other sugars. Distribution of sialic acid in organisms is limited as compared with glucose, which is universally distributed in organisms. The structural diversity and maldistribution of sialic acid may have arisen during the course of evolution. The origin of sialic acids may be traced back to by the maldistribution of sialic acid, and the history of sialic acids may be speculated on based on the structural diversity of sialic acid as well as the specificity of its binding counterpart molecules. |
|
|
|
|
Methods to detect sialic acids |
It is very important to verify the presence or absence of sialic acids in various organisms in the phylogenic tree (not only living organisms but extinct fossils as well). So far, many researchers have demonstrated the presence of sialic acid in various organisms. To demonstrate the presence of sialic acid, the sample is subjected to colorimetric analyses using the resorcinol (RCH) reagent or thiobarbituric acid (TBA) reagent and, for firm evidence, to gas-chromatography, mass-spectrometry, and NMR spectrometry. The detection limit of sialic acid by the RCH and TBA reagent is of the pmol order. Recently, the development of an α-keto acid specific fluorescent reagent, DMB (1,2-diamino-4,5-methylenedioxybenzene)4) together with the improvement of sensitivity of the fluorescent detector dramatically improved the detection limit of sialic acid to the fmol to amol order. This highly sensitive detection method enabled us to detect sialic acid even in fossil samples and to gain an insight into the evolution of sialic acids. However very careful procedures to remove contaminants including those from the researchers themselves, other samples, and the environment (food) are required.
Another way to see the presence of sialic acid is to examine the genes responsible for the biosynthesis of the acid. Recently, genome projects on several selected animals have ended and the databases have been rapidly compiled and wide available. In addition, most of the key enzymes responsible for the biosynthesis of sialic acid have been cloned. It is thus easy to speculate on the ability of organisms in question to produce sialic acids.
|
|
|
|
|
Demonstration of the presence of sialic acids--biochemical evidence |
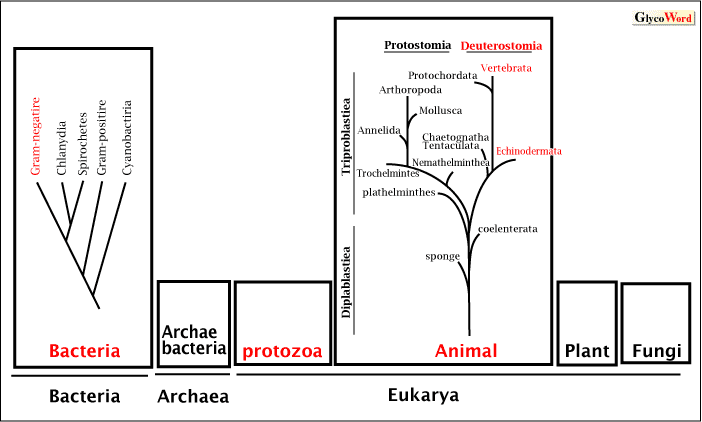
| Many researches showed the presence of sialic acids in various organisms (Fig.1). In prokaryotic cells, some gram-negative and pathogenic bacteria groups contain sialic acids, although the presence of sialic acid in archaebacteria has not been reported. In eukaryotes, i.e., the plant, fungal, animal (protostomia, and deuterostomia) and protozoa kingdom, reports of the presence of sialic acid were restricted in animals and protozoa especially in deuterostome. In practice, almost all the reports came from vertebrates and echinoderms. However, several reports have recently appeared indicating that plant (buckwheat and Arabidopsis thaliana), fungi (Asperigillus, Candida), Arthropoda (Dorosophila) and Mollusca (squid) that are categorized in protostomia have sialic acids. |
|
|
|
 |
|
Figure 1 Organisms and the occurrence of sialic acids
Organisms in red have been reported to have sialic acid.
|
| |
|
|
|
|
|
Speculation of the presence of sialic acids--evidence from bioinformatics on genomes |
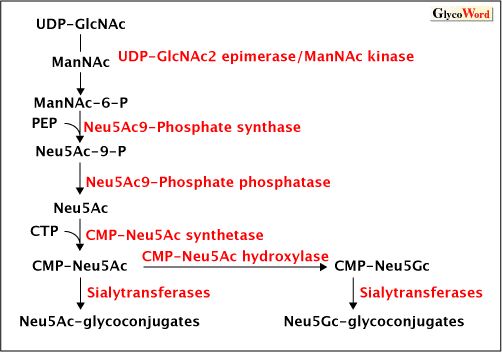
| Today, we can easily get various genomic information of various selected animals from databases of national projects on genomes. Although a glycan is a secondary gene product, we can speculate the presence of the sugar in organisms if we find the existence of key enzymes that are required during the biosynthesis of the sugar. The biosynthetic pathway of sialic acids is shown in Fig.2. The essential biosynthetic enzymes of sialic acids are UDP-GlcNAc 2-epimerase/ManNAc kinase, sialic acid 9-phosphate synthetase, sialic acid 9-phosphate phosphatase, sialic acid transporter, CMP-Sia synthetase, and sialyltransferases. The presence of the homologous genes that encode such key enzymes leads us to speculate the presence of a known biosynthetic pathway. Searching the genome database for such biosynthetic information, some conflicts are noted between the results from biochemical analyses and bioinformatics. A conclusion cannot be reached easily because the sialic acid may be biosynthesized via an as-yet unknown pathway or be taken up from the environment via a host-derived reaction such as the trans-sialidase activity of Trypanosoma (see trans-sialidase). Moreover, databases have not yet been perfected. |
|
| |
|
 |
|
Figure 2 Biosynthetic pathway of sialic acids
Enzymes in red are key enzymes. We can speculate on the occurrence of sialic acid by searching the database for the presence of these enzymes.
|
| |
|
|
Neu5Ac and Neu5Gc |
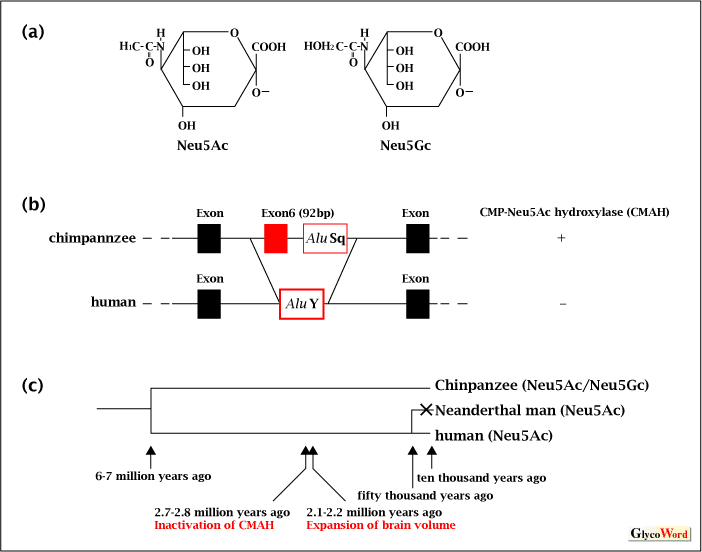
Due to the structural diversity of sialic acids, it is important to know the occurrence of sialic acid species in detail. Because of the diversity in its substitution, comprehensive analyses have not been reported. However, the comparative localization of Neu5Ac and Neu5Gc has been well studied (Fig.3(a)). Neu5Gc appears on the cell surface as follows. CMP-Neu5Ac, which is the donor of sialyltransferases, is converted to CMP-Neu5Gc by active-CMP-Neu5Ac hydroxylase6,7) to hydroxylate the acetyl-group at the C5 position (Fig.2). The CMP-Neu5Gc is then used as a donor substrate like CMP-Neu5Ac by sialyltransferases and Neu5Gc is transferred to the glycoconjugates. Not only Neu5Ac but also Neu5Gc are observed in the organisms that have active CMP-Neu5Ac hydroxylase. So far, human and bird are reported to have only Neu5Ac. Although human has exclusively Neu5Ac, chimpanzee and orangutan of the same primate family have both Neu5Ac and Neu5Gc (Fig.3(b))8). The reason why human does not have Neu5Gc is not that human does not have a CMP-Neu5Ac hydroxylase enzyme but that human has an inactivated (incomplete) CMP-Neu5Ac synthetase that can not hydroxylate CMP-Neu5Ac due to the insertion of the AluY sequence into the genome that encodes CMP-Neu5Ac hydroxylase followed by deletion of exon 6 (See N-Glycolylneuraminic acid and N- Acetylneuraminic acid)6,7,8)(Fig.3(c)).
Although the enzyme is inactivated, the occurrence of a very tiny amount of Neu5Gc has been reported in human. This is considered to come from the environment such as food or culture medium9). It is interesting to take into account the maldistribution of the Neu5Gc and Neu5Ac between the organisms when we consider evolution, especially in case of chimpanzee and human. These two animals are said to have split from a common ancestor 5-6 million years ago and have very highly homologous genes (homology- 97%). Only 3 % of the genes is different between the two species and it is noteworthy that CMP-Neu5Ac hydroxylase is one such genes. In addition, the insertion of the AluY sequence into human genome is considered to have occurred 2.7-2.8 million years ago. This era was just before the Stone Age and just before the age that the brain volume of human increased remarkably (2.1 million years ago). These phenomena may be related to the fact that human has exclusively Neu5Ac5,8). In addition, it is very interesting that all vertebrates that have both Neu5Ac and Neu5Gc in various tissues contain exclusively Neu5Ac in the brain. Such findings lead us to imagine that human might have selected Neu5Ac after using diverse sialic acids like Neu5Gc (via degeneration of the gene)5,8). |
|
|
| |
|
 |
|
Figure 3 Neu5Ac and Neu5Gc
(a) Structure of Neu5Ac and Neu5Gc
(b) Comparison of the genome which encodes CMP-Neu5Ac hydroxylase (CMAH) in chimpanzee and human
(c) Phylogenic tree of chimpanzee, Neanderthal man and human and the occurrence of sialic acid species detected.
|
| |
|
|
Lectins that make sialic acid more diverse |
| Lectins that recognize sialic acids are categorized into three groups, pathogenic lectins towards glycoconjugates from vertebrates, endogenous lectins in vertebrates, and exogenous lectins from other organisms such as insects and plants. Plants have lectins that recognize sialic acids and the plant lectins are powerful tools for the detection of sialic acids. Because plant itself does not have sialic acids, plant lectins are considered to function in protecting itself from foreign enemies that have sialoglycoconjugates. Alternatively, the real ligands for these lectins may be mimetic molecules different from sialic acids. Pathogenic lectins from virus, bacteria, and protozoa are involved in infection through their sialic acid-recognizing activity. One of the best studied examples is hemagglutinin of influenza (see influenza session) in which the virus utilizes sialic acids on the cell surface of the host during infection. The specificity of hemagglutinin changes depending on subtle structures of the hosts sialic acids (linkage and species). Diversity in sialic acids on the cell surface of the host may be the traces remaining as a result of the battles between host and pathogen. Getting rid of the sialic acid from itself is an easy way to escape the infectious pathogens that utilize host sialic acids, but this would be impossible because sialic acids might be too important to remove. The report that sialic acid-deficient mice are embryonic lethal is one piece of evidence that sialic acids are essential. Thus it is reasonable to imagine that the diversity in sialic acid is a trace of the battles that raged between pathogenic bacteria and host via changing linkage or substitution of the sialic acids. However, it is difficult to verify this. Diversity in sialic acid structure on the host cell may arise not only from exogenous sialic acids-recognizing lectins but also from endogenous sialic acids-recognizing lectins such as complement H, selectin (See selectin session) and siglec (See siglec session). The binding specificities of these lectins are different according to the sialic acid species and linkages, and may be the result of frequent encounters with endogenous lectins. In brief, the more diverse the sialic acid structure becomes by species, substitution of the hydroxyl group with sulfation, or methylation by adding more sialic acids toward sialic acids and by changing the structure of inner glycan chains, the more liable the endogenous lectins are to be multifunctional and beneficial to the organism. |
|
|
| |
|
|
Origin of sialic acids |
Three possibilities for the origin of sialic acid are proposed3). First, sialic acids developed before the three branches (bacteria, archae, eukaryotes) divided so present in each. Second, bacteria developed sialic acids, and the genes for enzymes involved in the biosynthesis of sialic acids transferred horizontally to archaea and eukaryotes. Third, eukaryotes developed sialic acids, and the genes for enzymes involved in the biosynthesis of sialic acid transferred horizontally to bacteria.
Sialic acids were first introduced to various organisms and the lectins that recognize (utilize) sialic acids were acquired. Then the sialic acids and those exogenous and endogenous lectins interacted with each other. The history of co-evolution under pressure from these interactions may account for the diversity in sialic acids and diversity in the specificity of sialic acid-recognizing molecules (lectins). |
|
|
|
|
Chihiro Sato (Bioscience and Biotechnology Center, Nagoya University) |
|
|
|
|
|
| References |
(1) |
Blix FG, Gottschalk A, Klenk E: Proposed nomenclature in the field of neuraminic and sialic acids. Nature, 179, 1088, 1957
|
|
(2) |
Nadano D, Iwasaki M, Endo S, Kitajima K, Inoue S, Inoue Y: A naturally occurring deaminated neuraminic acid, 3-deoxy-D-glycero-D-galacto-nonulosonic acid (KDN). Its unique occurrence at the nonreducing ends of oligosialyl chains in polysialoglycoprotein of rainbow trout eggs. J Biol. Chem., 261, 11550-11557, 1986
|
|
(3) |
Angata T, Varki A: Chemical diversity in the sialic acids and related alpha-keto acids:an evolutionary perspective. Chem. Rev., 102, 439-469, 2002 |
|
(4) |
Hara S, Takemori Y, Yamaguchi M, Nakamura M, Ohkura Y: Fluorometric high-performance liquid chromatography of N-acetyl- and N-glycolylneuraminic acids and its application to their microdetermination in human and animal sera, glycoproteins, and glycolipids. Anal Biochem., 164, 138-145, 1987 |
|
(5) |
Chou HH, Hayakawa T, Diaz S, Krings M, Indriati E, Leakey M, Paabo S, Satta Y, Takahata N, Varki A: Inactivation of CMP-N-acetylneuraminic acid hydroxylase occurred prior to brain expansion during human evolution. Proc. Natl. Acad. Sci. USA, 99, 11736-11741, 2002 |
|
(6) |
Irie A, Koyama S, Kozutsumi Y, Kawasaki T, Suzuki A: The molecular basis for the absence of N-glycolylneuraminic acid in humans. J. Biol. Chem., 273, 15866-15871, 1998 |
|
(7) |
Chou HH, Takematsu H, Diaz S, Iber J, Nickerson E, Wright KL, Muchmore EA, Nelson DL, Warren ST, Varki A: A mutation in human CMP-sialic acid hydroxylase occurred after the Homo-Pan divergence. Proc. Natl. Acad. Sci. USA, 95. 11751-11756, 1998 |
|
(8) |
Hayakawa T, Satta Y, Gagneux P, Varki A, Takahata N: Alu-mediated inactivation of the human CMP-N-acetylneuraminic acid hydroxylase gene. Proc. Natl. Acad. Sci. USA, 98, 11399-11404, 2001 |
|
(9) |
Tangvoranuntakul P, Gagneux P, Diaz S, Bardor M, Varki N, Varki A, Muchmore E: Human uptake and incorporation of an immunogenic nonhuman dietary sialic acid. Proc. Natl. Acad. Sci. USA, 100, 12045-12050, 2003 |
|
(10) |
Schwarzkopf M, Knobeloch KP, Rohde E, Hinderlich S, Wiechens N, Lucka L, Horak I, Reutter W, Horstkorte R: Sialylation is essential for early development in mice. Proc. Natl. Acad. Sci. USA, 99, 5267-5270, 2002 |
|
(11) |
Varki A. in Essentials of glycobilogy (Varki A, Cumminings R, Esko J, Freeze H, Hart G. and Marth J. eds), pp195-209, 1999 |
|
|
|
|
|
|
|
|
| Sep. 29, 2006 |
|
|
|
|
|
|
|



