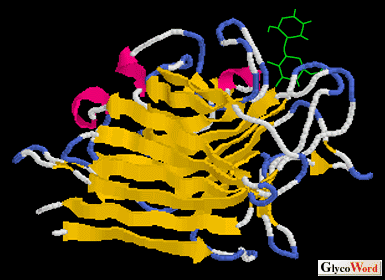
| Legume lectins are one of the largest lectin families with more than 70 lectins were reported. Leguminous plant lectins resemble each other in their physicochemical properties although they differ in their carbohydrate specificities. They consist of two or four subunits with relative molecular mass of 30 kDa and each subunit has one carbohydrate-binding site. The interaction with sugars requires tightly bound calcium and manganese ions. The primary structural analyses and X-ray crystallographic studies report the structural similarities of these lectins. X-ray studies have shown that the folding of the polypeptide chains in the region of the carbohydrate-binding sites is also similar, despite differences in the primary sequences. The carbohydrate-binding sites of these lectins consist of two conserved amino acids on beta pleated sheets and two loops, as shown in the figure. One of these loops contains transition metals, calcium and manganese, and keep the amino acid residues of the sugar binding site at the required positions. Amino acid sequences of these loops play an important role in the carbohydrate-binding specificities of these lectins.
The most likely function for the plant lectins is plant defense. Especially in legume lectins, they are also involved in the recognition of bacteria for the purpose of establishing symbiosis. Proteins with leguminous lectin domains were found in animal cells. Cargo receptors, ERGIC-53 and VIP36, have a leguminous lectin domain on their lumenal side and localize to the cis Golgi network and trans Golgi network, respectively. They operate sorting and transport of newly synthesized glycoproteins from ER through Golgi to the membrane. | |
|
| References | (1) | K. Yamamoto.: Structure and function of lectins and lectin-carbohydrate interactions. Seikagaku 66, 1111-1129, 1994 |
| (2) | Yamamoto, K, Konami, Y, Osawa, T: Purification and characterization of a carbohydrate-binding peptide from Bauhinia purpurea lectin. FEBS Lett. 281, 258-262, 1991 |
| (3) | Yamamoto, K, Konami, Y, Osawa, T, Irimura, T: Carbohydrate-binding peptides from several anti-H(O) lectins. J. Biochem. 111, 436-439, 1992 |
|
(4) |
Yamamoto, K, Konami, Y, Osawa, T, Irimura, T: Alteration of carbohydrate-binding specificity of Bauhinia purpurea lectin through the construction of chimeric lectin. J. Biochem. 111, 87-90, 1992 |
|
(5) |
K. Yamamoto, Y. Konami, T. Osawa.: Chimeric lectin of Bauhinia purpurea lectin and Lens culinaris lectin recognizes unique carbohydrate structure. J. Biochem. 127, 129-135, 2000 |
|
(6) |
K. Yamamoto, I. N. Maruyama, T. Osawa.: Cyborg lectins: Novel leguminous lectins having unique specificities. J. Biochem. 127, 137-142, 2000 |
| | |
| |