|
|
カブトガニレクチンの糖鎖認識機構 |
 |
||||||||||
 |
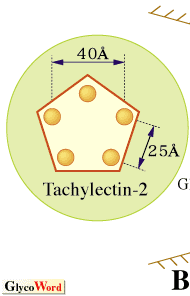
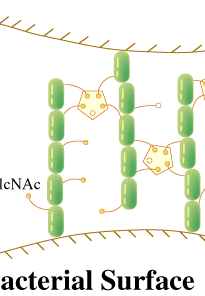
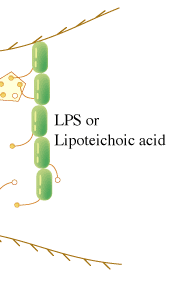
カブトガニ血球および血漿からタキレクチンと名づけられた5種類のレクチンが精製された(1)。タキレクチン-1は、グラム陰性菌由来のリポ多糖(LPS)に共通の構成糖である2-ケト-3-デオキシオクトン酸(KDO)に結合することでグラム陰性菌を認識し、また、アガロースやデキストランに吸着する性質がある。タキレクチン-2は、生物に普遍的にみられるD-GlcNAcやD-GalNAcを結合し、ある種のグラム陽性菌由来のリポテイコ酸やLPSを認識する。一方で、タキレクチン-3やタキレクチン-4は、特定のLPSのO-抗原(S-型LPS)に見られる糖や糖鎖構造を特異的に識別することができる。血漿中に存在するタキレクチン-5は、これまで調べられたカブトガニレクチンの中では、最も強い凝集活性を示すが、その認識の標的は、より構造的に単純なアセチル基である(2)。タキレクチン-5は、N-末端のCys含有領域と哺乳類のフィコリンと高い配列類似性を示すフィブリノーゲンドメインからなっている。タキレクチン-5は、2枚から4枚羽根のプロペラ構造をしていることが電子顕微鏡観察から明らかになった。この構造は、フィコリンやコレクチンの花束構造と類似しているが、コラーゲンドメインを欠いている。カブトガニの体液には、上記のような特異性の大きく異なった生体防御レクチンが備わっており、傷を受けた局所において、これらのレクチンが協調的に機能していると考えられる。 最近、タキレクチン-2のX線結晶構造が決定された(3)。タキレクチン-2の立体構造は、一次構造の5回繰り返し配列を反映して、5つのβ-プロペラ構造から構成されている。さらに、レクチン-リガンド複合体の構造解析から、それぞれのプロペラに1個のD-GlcNAcが結合していることが判明した。従って、生体防御レクチンの非自己認識のひとつの方法は、感染微生物特異的な物質を認識するのではなく、生物に普遍的に見い出される糖鎖リガンドの高い表面密度を認識することに他ならない(図参照)。個々の結合部位とD-GlcNAcとの親和性は安定な結合を維持するほど強くはないが、結合部位を多価にして親和性を増強させるとともに、それぞれの結合部位を近接させることにより、D-GlcNAcが高密度で存在している状態、すなわち感染微生物表面の分子パターンに対する非自己認識の高い特異性が生み出されている。 |
||||||||||
|
|
|||||||||||
|
|||||||||||
| 川畑俊一郎(九州大学大学院理学研究科) | |||||||||||
|
|||||||||||
| 2000年 12月 15日 | |||||||||||
|
|
|||||||||||
|
|||||||||||