|
|
Protein Engineering of the Reaction Mechanism of Cyclodextrin Glucanotransferase |
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
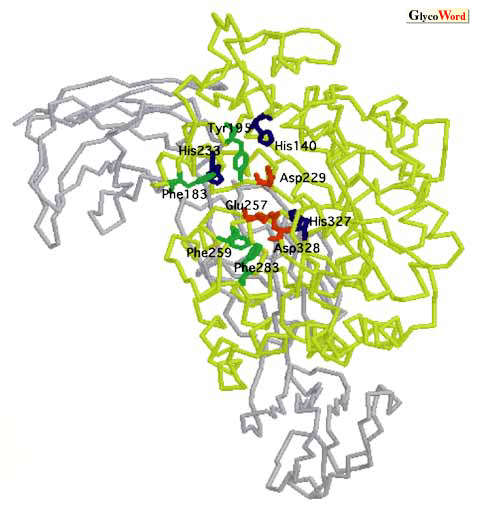
Cyclodextrin glucanotransferases (CGTase EC 2.4.1.19) are amylolytic enzymes derived from bacterial sources that catalyze the conversion of starch into a mixture of cyclodextrins (CDs). In general, CDs consist of six, seven or eight glucose units (alpha-, beta- or gamma-CDs) through an intramolecular transglycosylation reaction. The known CGTases are classified as alpha-, beta- or gamma-CGTases based on their main CD product. Moreover, CGTases catalyze two types of intermolecular transglycosylation reactions, i.e. coupling and disproportionation, and slightly catalyze hydrolysis. Cyclodextrins can form inclusion complexes with a variety of molecules and alter the chemical and physical properties of the included compounds. Therefore, CGTases are employed for example, in the food, cosmetic and pharmaceutical industries. Protein engineering has been combined with the alignment of numerous sequences, mutational analysis and three-dimensional structures of intact and mutant CGTases complexed with substrate analogues and natural substrate. This has allowed some functional amino acid residues in the CGTase reaction to be elucidated. Essential residues of catalysis The CGTases are functionally related to alpha-amylase (EC 3.2.1.1) (Family13), having sequence similarity (30%) and clear structural similarity of the N-terminal A-C domains: catalytic A domain folds into an (beta/alpha)8-barrel structure and the B, C, and two additional D-E domains consist mainly of a loop region and beta-sheets. Kinetic studies on the CGTase from Bacillus circulans strain 8 and the X-ray structure of CGTase from B. circulans strain 251 ligated to the oligosaccharide analogue, have revealed a total of nine subsites (-7 to 2) (1,2). The three carboxylates that are highly conserved in family 13, Asp229, Glu257 and Asp328 (alkalophilic Bacillus sp.#1011 CGTase numbering) may be catalytic residues (3) because the enzymatic properties after replacing the acids with the corresponding amides show that the CD-forming activity (4) of each mutant significantly decreases (4,000-700,000 folds). Three histidine residues (His140, His233, and His327) are located at the active center of the enzymes in family 13. Mutational analysis of these residues using Asn suggests that they stabilize the transition state through hydrogen-bonding contacts (5). X-ray structural studies of CGTase substrate or substrate analogue complexes have revealed detailed interaction between ligands and key functional amino acid residues during CGTase catalysis, as well as alpha-amylase family (Family 13) catalysis (2, 6-9). (Fig. 1) In the proposed catalytic mechanism of CGTase, Glu257 plays the acid/base catalyst, Asp229 the catalytic nucleophile and Asp328 stabilizes substrate binding and elevates the pKa of Glu257. (See details on SA-B03.) His140 and His327 are in close contact with Glu257, Asp229 and Asp328, and they undergo distortion from the chair conformation of the sugar residue in subsite -1 (transition state) and the covalent bond between Asp229 and the C1 of the sugar residue in subsite -1 in an intermediary complex. Moreover, the charged residues described above, undergo catalysis with the help of Tyr100 (stacking interaction) and Arg227 (hydrogen bonds). |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Essential residues of cyclization
Comparisons of the primary and tertiary structures of CGTases with alpha-amylases have revealed that the four aromatic amino acids, Phe183, Tyr195, Phe259 and Phe283, commonly located in the active site of CGTase (Fig. 2) are not located in that of alpha-amylases., which consist almost totally of aliphatic hydrophobic residues. Y195L-CGTases (10, 11) and Y195W-CGTase (12) affect the size of the CD produced, namely the larger beta- and gamma-CDs. Structural data of H233N-CGTase complexed with acarbose, a maltotetraose analogue, indicate that the missing interactions between His233 and the sugar residue in subsite 1 caused the weak affinity of acarbose on subsite 1, resulting in significantly decreased alpha-CD production (13). F183L/F259L-CGTase considerably decreases cyclization efficiency, although the intermolecular transglucosylation activity is very similar to that of wild-type CGTase (10). F255I-CGTase from B.stearothermophilus (Phe255 is equivalent to Phe259) also has diminished CD-forming activity (14). Thus, the stacking interaction with both Phe183 and Phe259 plays a critical role in cyclization when the nonreducing end of amylose binds to the active center of CGTases. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
On the other hand, replacing Phe283 located near the acid/base catalyst (Glu257) with a leucine residue significantly changes the pH profile of starch-degrading activity (10). Hydrophobic residues are constantly located around the Glu257 in alpha-amylases and CGTases (7, 15-18). The three-dimensional structure of Taka-amylase A (TAA) suggests that the pKa value for Glu230 is increased by the hydrophobic environment (18, 19). These findings indicate that Phe283 is important for the hydrophobic environment around Glu257, and hence participates in raising the pKa value for Glu257 (11).
Structural analysis of the inactive mutant E257A-CGTase from B.circulans strain 8 soaked in the CD derivative S-(beta-D-glucopyranosyl)-6-thio-alpha-CD (thioglucosyl-CD) indicates a state immediately after cyclization or before coupling in the reverse reaction. However, details of the interaction between this ligand and the residues constituting each subsite remain unclear (20). Further studies aimed at changing the properties of CGTases should be performed based the current insight into structure/function relationships. Such studies could result in the design of a CGTase to produce a single type of CD. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Keiko Haga and Kunio Yamane (Institute of Biological Sciences, University of Tsukuba) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mar. 15, 2000 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||