|
|
サイクロデキストリン合成酵素の反応機構の蛋白質工学的解析 |
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
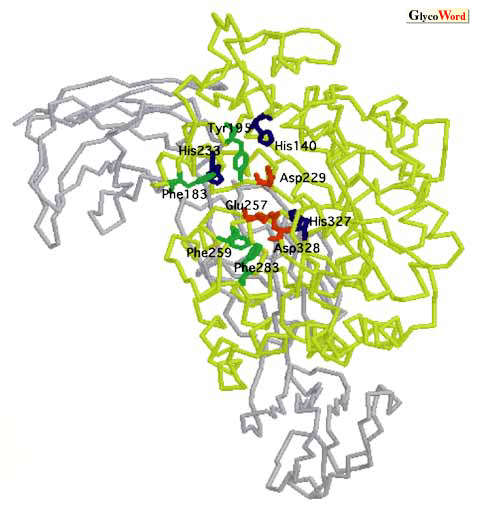
サイクロデキストリン合成酵素(EC 2.4.1.19)は細菌起源の糖質分解酵素で、デンプンからサイクロデキストリンを生成する反応を触媒する酵素である。 分子内転移反応によって生成されるサイクロデキストリンはグルコースが6個、7個、あるいは8個単位から構成されるα- サイクロデキストリン、β-サイクロデキストリン、γ-サイクロデキストリンが一般的である。 これまでに報告されているサイクロデキストリン合成酵素は主に生成するサイクロデキストリンによって、 α- 、β-、あるいはγ-サイクロデキストリン合成酵素に分類されている。 このほかに、サイクロデキストリン合成酵素は2つの分子間転移反応であるカップリング反応や不均化反応、さらには若干の加水分解反応も触媒する。 サイクロデキストリンは様々な分子を包み込む包接化合物を形成しそれらの化学的・物理的性質を変化させることが可能であるため、サイクロデキストリン合成酵素は食品、薬品、化粧品産業へ応用利用されている。 サイクロデキストリン合成酵素の反応機構の蛋白質工学的解析は、膨大な数の一次配列の比較や部位特異的変異による解析、あるいは野生型酵素と変異酵素の天然基質や基質類似体との複合体の3次構造の解析を組み合わせておこなわれいるが、これらの解析によってサイクロデキストリン合成酵素の反応に重要な役割を持ついくつかのアミノ酸が解明されている。 触媒反応に不可欠なアミノ酸残基 サイクロデキストリン合成酵素は α-アミラーゼ(EC 3.2.1.1)(ファミリー13)と機能的に関連のある酵素で、アミノ酸の一次配列の相同性は30%であるが、N末端側のA-Cドメインには非常にはっきりとした構造的類似性がある: 触媒ドメインである Aドメインは(β/α)8バレルという構造をとり、B、Cドメインとさらに付加されたD、Eドメインはおもにループやβシートから構成されている。Bacillus circulans strain 8 由来サイクロデキストリン合成酵素のキネティクスのデータやオリゴ糖の類似体が結合したB. circulans strain 251 由来サイクロデキストリン合成酵素の三次構造の解析結果から、サイクロデキストリン合成酵素には9つ(-7から2)のサブサイトが存在することが明らかになっている(1, 2)。 アスパラギン酸229、グルタミン酸257及びアスパラギン酸328の3つのカルボン酸はファミリー13に属する酵素間で高度に保存されており(アミノ酸残基の番号はBacillus sp.#1011 CGTaseに準ずる)触媒残基と推定されているが(3)、これらのアミノ酸をそれぞれのアミドに置換した変異体酵素のサイクロデキストリン生成活性は野生型酵素の4,000から700,000分の1と著しく減少している(4)。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3つのヒスチジン残基、 ヒスチジン140,ヒスチジン233、 およびヒスチジン327はファミリー13に属する酵素の活性中心に存在している。 これらのヒスチジンをアスパラギンに置換した変異体酵素の解析結果からこれらのヒスチジン残基が水素結合を介して遷移状態の安定化に寄与していることが推測されている(5)。 サイクロデキストリン合成酵素と基質および基質類似体との複合体のX線解析の研究は、サイクロデキストリン合成酵素の触媒反応時における、結合した基質や基質類似体と反応に重要に関わっているアミノ酸との間の相互作用の詳細な解明につながり、これらはサイクロデキストリン合成酵素のみならず、α-アミラーゼファミリー(ファミリー13)に属する酵素全般の触媒反応において適用されると報告している(2, 6-9)。 提唱されたサイクロデキストリン合成酵素の反応機構において、グルタミン酸257は酸/塩基触媒として、アスパラギン酸229は求核剤として働き、アスパラギン酸328は基質との結合の安定化とグルタミン酸257のpKa値の引き上げに関与していると考えられる。(詳細はSA-B03を参照)この際、ヒスチジン140とヒスチジン327はグルタミン酸257、アスパラギン酸229およびアスパラギン酸328と密接に相互作用してサブサイト-1に結合した基質の糖残基の椅子型構造を歪ませ(遷移状態)、さらに中間複合体におけるアスパラギン酸229とサブサイト-1の糖残基のC1間の共有結合の形成を担っている。 さらに、これらの荷電アミノ酸はチロシン100(スタッキング)やアルギニン227(水素結合)の助けを借りて触媒反応を遂行している。 環形成反応に不可欠なアミノ酸残基 サイクロデキストリン合成酵素とα-アミラーゼの一次構造や三次構造を比較するとサイクロデキストリン合成酵素の活性中心には4つの芳香族アミノ酸、フェニルアラニン183、チロシン195, フェニルアラニン283、およびフェニルアラニン259が保存されている(Fig. 2)。しかし、α-アミラーゼの活性中心に存在しているのはほとんどが脂肪族の疎水性アミノ酸である。チロシン195をロイシンに置換した変異体酵素(10, 11)、Y195L-CGTaseとトリプトファンに置換した変異体酵素、Y195W-CGTase(12)の生成するサイクロデキストリンは、β-サイクロデキストリンやγ-サイクロデキストリンといったより大きいサイズのサイクロデキストリンに変化する。 ヒスチジン233をアスパラギンに置換した変異体酵素(H233N-CGTase)とマルトテトラオースの類似体である、アカボースとの複合体のX線解析は、ヒスチジン233とサブサイト1に結合する糖残基との相互作用の欠如がサブサイト1のアカボースに対する親和性を弱め、その結果、 α-サイクロデキストリン生成活性の減少を引き起こす事を示唆している(13)。 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| フェニルアラニン183とフェニルアラニン259をロイシンに置換した二重変異酵素、F183L/F259L-CGTaseは環形成反応の効率が極端に減少するが、分子間転移活性は野生型酵素とほぼ同じである(10)。 B. stearothermophilus 由来のサイクロデキストリン合成酵素ではフェニルアラニン259はフェニルアラニン255に相当するが、これをイソロイシンに置換してもやはり、環形成活性は減少する(14)。従って、環形成反応においてアミロースの非還元末端がサイクロデキストリン合成酵素の活性中心に結合する時、フェニルアラニン183とフェニルアラニン259の双方のスタッキングが非常に重要になるといえる。一方、酸/塩基触媒であるグルタミン酸257の近傍に存在するフェニルアラニン283 をロイシンに置換するとデンプン分解活性のpH依存性が著しく変化する(10)。 α-アミラーゼやサイクロデキストリン合成酵素ではグルタミン酸257の近傍に疎水性アミノ酸残基が広く存在している(7, 15-18)。タカアミラーゼAの構造解析によりグルタミン酸230(グルタミン酸257に相当)の pKa値は周囲の疎水的な環境によって引き上げられることが推定されている(18, 19 )。以上の知見からもフェニルアラニン283はグルタミン酸257の近傍の疎水的環境を形成しそのpKa値の引き上げの役割を担っているものと思われる。 B.circulans strain 8 由来のサイクロデキストリン合成酵素において活性のない変異酵素、すなわちグルタミン酸257をアラニンに置換した酵素(E257A-CGTase)にサイクロデキストリンの誘導体S-(β-D-glucopyranosyl)-6-thio-α-CD (チオグルコシルサイクロデキストリン)をソーキングした解析結果は、環形成直後の状態かその逆反応のカップリングの直前の状態を示唆しているが、それぞれのサブサイトを構成するアミノ酸残基と結合した基質間の相互作用の詳細な情報は得られていない(20)。 これまで得られたサイクロデキストリン合成酵素の構造と機能に関する知見をもとにして、今後もサイクロデキストリン合成酵素の特性を変えるような研究が行われるべきであり、結果として単一のサイクロデキストリンを生成する酵素の構築も可能になると思われる。 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 羽賀敬子・山根國男(筑波大学 生物科学系) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2000年 3月 15日 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||