
Haruko Shirato-Horikoshi
Department of Virology II, National Institute of Infectious Diseases.
Haruko Shirato graduated from the Faculty of Veterinary Medicine, Hokkaido University in 1997, and received her Ph.D. degree at the Graduate School of Medicine, Osaka University in 2001. She was appointed to research associate from 2001 to 2007 at the Department of Virology II at the National Institute of Infectious Diseases, and is currently senior research associate since 2007. Her recent research interest is viral infection mechanism controlled by carbohydrates.
Viruses initiate infection by attaching to specific cells of target host tissue. Virus receptors are greatly involved in host-, tissue-, and cell-specificity, and carbohydrate molecules are among those used for virus attachment. Orthomyxoviruses, polyomaviruses, reoviruses, coronaviruses, paramyxoviruses, and both murine and canine parvoviruses use sialic acid for binding, whereas adenovirus-associated virus 2, herpes viruses, and flaviviruses recognize heparan sulfate for binding1. Recently it was found that norovirus (NoV) appears to recognize and bind to histo-blood group antigens (HBGAs), which are uncharged sugar residues. Here we summarize the interaction between NoV and HBGAs.
NoV's clinical features, infection mechanisms, virological classification, and characteristics are summarized.
NoV is the major causative agent of acute viral gastroenteritis worldwide. Excluding cases caused by bacteria (Salmonella, Staphylococcus, Vibrio, Campylobacter, Escherichia coli, Clostridium perfringens), NoV accounts for more than 95% of virus-borne food poisonings, and shows similar distribution patterns every year (Fig. 1). NoV is also the main causative virus of infectious gastroenteritis prevalent in winter. Although NoV usually causes a short-term, self-limiting disease, oral rehydration and intravenous replacement of electrolytes are needed when severe diarrhea is observed. Complications from NoV infection are usually observed in infants and the elderly because they are generally more sensitive to water deprivation and tend to suffer aspiration pneumonia.
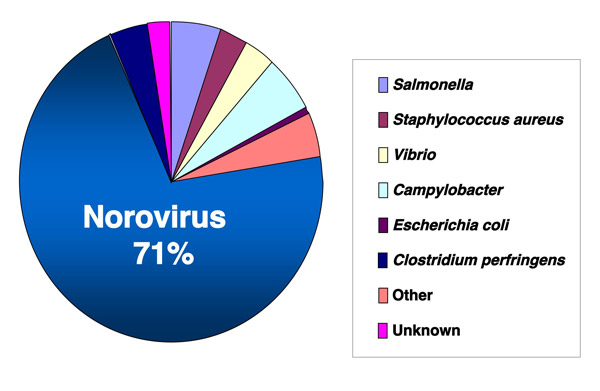
Fig. 1 Proportion of food poisonings by bacteria or virus in 2006, Japan.
The majority (27,642 of 39,222, or 71%) of cases involved diarrhea due to NoV infection. Data are based on the reports of Food Poisoning Statistics of the Ministry of Health, Labor, and Welfare as of May 2007.
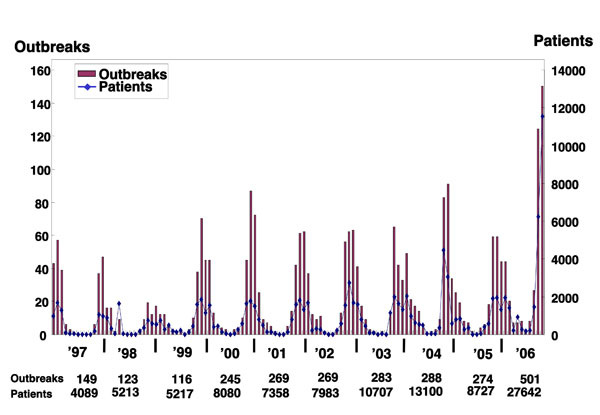
Fig. 2 The numbers of outbreaks and patients, per month, due to NoVs.
Based on a report from the Ministry of Health, Labor, and Welfare Food Poisoning Statistics as of May 2007.
As NoV infection occurs only in humans, and since no cell culture system has been developed yet, molecular analysis of the mechanisms underlying infection or reproductive replication has not advanced. The site of primary replication of NoV has not been established, but it is assumed to be the upper intestinal tract. Biopsies of the jejunum of volunteers who developed gastrointestinal illness following oral administration of NoV exhibit histopathologic lesions 2, where blunting of the villi of the proximal small intestine is observed.
NoV is classified as the type species Norwalk virus of the genus Norovirus in the family Caliciviridae (Table 1). There are four genera of Caliciviridae viruses-Vesivirus, Lagovirus, Norovirus, and Sapovirus-forming small, positive-stranded RNA viruses. Rabbit Hemorrhagic Disease Virus in the genus Lagovirus was the first calicivirus shown to bind to the HBGAs 3, prompting studies of the interaction between NoV and HBGAs.
Table. 1 Viruses included in the four genera of the family Caliciviridae
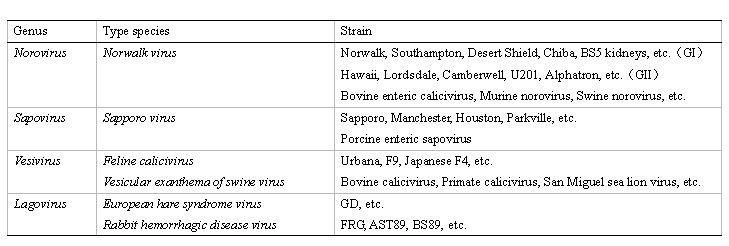 Most noroviruses and sapoviruses are isolated from humans. Recently, closely related viruses have been isolated from bovine, swine, and rodent, but there is no report indicating that these viruses are transmitted to humans.
Most noroviruses and sapoviruses are isolated from humans. Recently, closely related viruses have been isolated from bovine, swine, and rodent, but there is no report indicating that these viruses are transmitted to humans.
NoV contains a single-stranded positive-sense 7.6 kb RNA genome encoding three open reading frames (ORFs). ORF1 encodes a nonstructural polyprotein, and ORF2 and ORF3 encode the major capsid protein VP1 and minor capsid protein VP2, respectively (Fig. 3) 4. One virus particle is composed of 180 copies of VP1, a copy of the genome RNA, and a few copies of VP2 5,6. The virion is morphologically observed as a small, round, non-enveloped virus with a diameter of 38nm (Fig. 4). Because NoV is a non-enveloped virus, the capsid protein itself seems to have lectin-like activity and recognizes HBGAs.
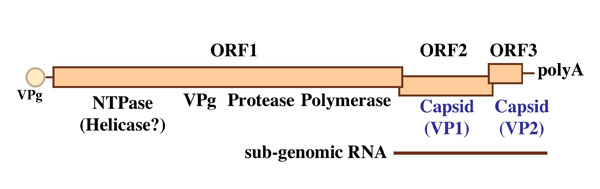
Fig. 3 Genome organization of NoV.
The genome encodes three ORFs. ORF1 encodes nonstructural proteins, ORF2 encodes capsid protein VP1, and ORF3 encodes minor capsid protein VP2.
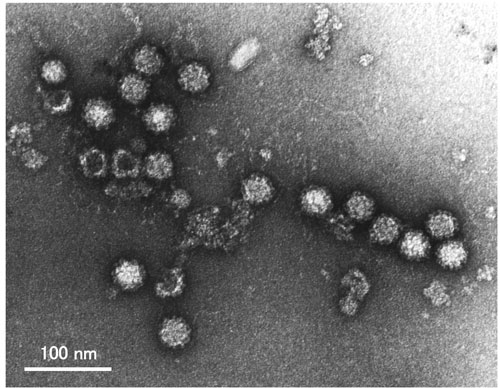
Fig. 4 Norovirus GI/4 Chiba407 strain visualized by electron microscopy.
The particle is observed as a small, round, non-enveloped virus with a diameter of 38nm.
Molecular biological studies of NoV began with the successful production of virus-like particles (VLPs). When VP1 or both VP1 and VP2 were expressed in insect or mammalian cells, a capsid protein of approximately 58 kDa appeared and self-assembled into VLPs 4,7. These artificial VLPs are morphologically and antigenically similar to the native virions. Many studies with VLPs have led to the development of sensitive solid phase immunoassays, including enzyme-linked immunosorbent assays (ELISAs) to detect the antigen. The expression of the VLPs also led to the identification of a cellular receptor or binding molecule for NoV.
NoV forms many morphologically similar but antigenically distinct groups of viruses 9. Recent progress in genetic studies has enabled us to divide human NoV into at least two genogroups - genogroup I (GI) and genogroup II (GII) - which contain at least 15 and 18 genotypes, respectively 8 (Fig. 5). This diversity makes the binding mechanism between NoV and HBGAs difficult to elucidate.
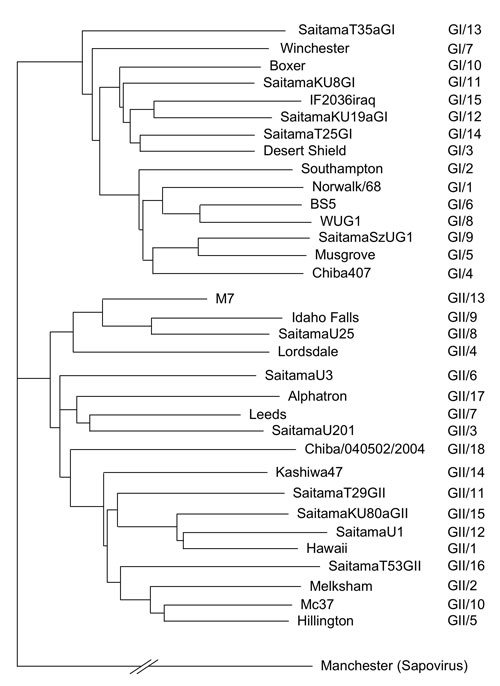
Fig. 5 Phylogenetic dendrograms based on the ORF2 gene of NoV.
Phylogenetic dendrograms are generated separately for GI (upper part) and GII (lower part). A representative strain of each genotype is shown. The clustering of genotypes is based on Ref 8. Manchester is used as the out group.
HBGAs are carbohydrates that contain structurally related saccharide moieties, ABH antigens, and Lewis antigens, etc. They are found in saliva and mucosal secretions from intestinal epithelial cells of secretor individuals who have the FUT2 gene encoding a fucosyltransferase. Non-secretors do not express FUT2 fucosyltransferase, and consequently do not express H type 1 or Le-b in the gut and saliva. For further information regarding ABH, Lewis antigens, and secretors/non-secretors, refer to Glycoprotein-B02 and A04 at this website.
The VLPs derived from the prototype strain of NoV, Norwalk virus (NV/68), have been shown to bind to H, A, and Le-b antigens by VLP-carbohydrate binding assay (Tables 2 and 3) 10-15. In the initial report, VLPs bound to the tissue sections. Thereafter, almost all reports used ELISA, the saliva-VLP binding assay, or the carbohydrate-VLP binding assay, and the attachment of the VLPs to HBGAs was detected and quantified.
Volunteer challenge studies provided strong evidence that carbohydrate binding is essential for NV/68 infection. Non-secretors were not infected after the challenge with NV/68. Furthermore, type O secretors were more likely to be infected with NV/68 and, conversely, type B secretors were less likely to be infected with NV/68 14,16. A report by Lindesmith et al. 14 had particularly great impact, because it included a large-scale infection experiment involving the examination of 77 volunteers. All of the infected individuals appeared to be secretors and, conversely, none of the non-secretors were infected.
NV/68 recognizes H, A, and Le-b antigens, as described above. However, other NoV VLPs display different ABH and Lewis carbohydrate-binding profiles, and indeed in vitro binding studies and epidemiological studies showed that some NoV strains could infect individuals with different ABH or secretor phenotypes 10-12, 17, 18. Therefore, each genotype seems to recognize different HBGAs. For example, the VLPs from BUDS strain bind A and B antigens, but not H antigen, whereas the VLPs from OIF strain bind only Le-a antigen (Tables 2 and 3).
Table. 2 Structure of synthetic carbohydrates
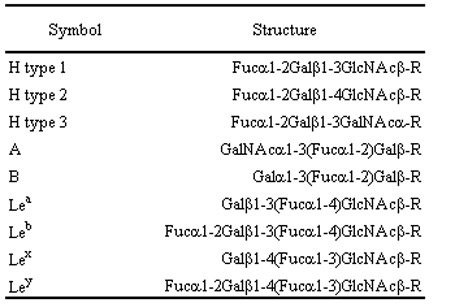 Glc, glucose; Fuc, fucose; Gal, galactose; GlcNAc, N-acetylglucosamine; GalNAc, N-acetylgalactosamine.
Glc, glucose; Fuc, fucose; Gal, galactose; GlcNAc, N-acetylglucosamine; GalNAc, N-acetylgalactosamine.
Table. 3 Carbohydrate binding patterns and genotype-specific residues of the putative binding sites
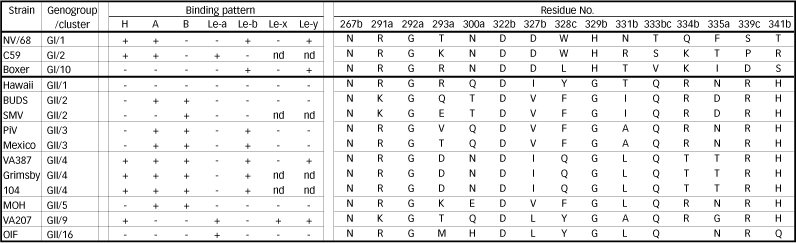
a; mutagenesis analysis (Ref 21)
b; evolution trace analysis (Ref 22)
c; X-ray crystallographic analysis (Ref 23)
The binding between HBGAs and GII/4 has been studied more extensively than other genotypes, and it has been shown that GII/4 VLPs bind to HBGAs more broadly and strongly than VLPs from other strains 11, 12, 19 (Tables 2 and 3). GII/4 is known to be a global epidemic genotype. In Japan, NoV outbreaks occurred in 236 healthcare facilities for the elderly during the winter of 2004–2005, causing 12 deaths in 7 facilities. Three NoV strains associated with three fatal cases were isolated from geographically separate facilities and genetically analyzed. The results revealed that all three isolates belonged to GII/4 20. It is hypothesized that the wide HBGA recognition of GII/4 strains may be linked with their strength of transmission.
Putative binding sites on the NoV capsid protein were identified by mutagenesis analyses and computer modeling 21, evolution trace analysis 22, and cocrystallization and X-ray analyses of recombinant P proteins with synthetic type A or B trisaccharides 23. The residues of putative binding sites are shown in Table 3. Strains of the same genetic cluster tend to have identical amino acids on putative binding sites. They are also known to have the same HBGA binding patterns. These observations are compatible with each other. The structural study of the P dimer-trisaccharide complex offered insight into the molecular aspect of their interaction. However, an entire virion consisting of 180 copies of the capsid protein and HBGAs, which are more complex than trisaccharides, are in vivo constituents. Therefore, it still remains to be elucidated whether the crystal structure of the P dimer-trisaccharide complex reflects the real phenomenon.
The recognition sites on HBGA by the prototype NV/68 strain and the GII/4 VA387 strain have been analyzed in detail. The attachment of NV/68 VLPs that bind to H, A, and Le-b antigens depends on the presence of α1,2 fucose (α-Fuc). This conclusion is based on the following observations: 1) NV/68 VLPs had no binding activity to H antigen precursors that lack the α-Fuc 10, 15; and 2) α1,2fucosidase treatment abolished the attachment of NV/68 VLPs to surface epithelial cells of the gastroduodenal junction as well as to saliva 15. On the other hand, the attachment of VA387 VLPs, which bind to H, A, B, Le-b, and Le-y antigens, depends on the presence of H (α-Fuc), A (N-acetylgalactosamine: α-GalNAc), or B (galactose: α-Gal) epitopes. This conclusion is based on the following observations: 1) the attachments of VA387 VLPs to A and B antigens were removed by α-N-acetylgalactosaminidase or a-galactosidase treatment 11; and 2) crystallization studies have suggested that α-Fuc, α-GalNAc, and α-Gal interact with the virus capsid protein 23. It remains to be determined whether or not NoV VLPs distinguish the difference between types 1, 2, and 3 carbohydrates. Since NoV forms many antigenically diverse groups (see paragraphs 2-6 and 3-4), and each genotype may recognize different HBGAs, the finding of the common NoV binding epitopes on host cells should be useful for the development of possible antiviral agents, and will help in the understanding of NoV transmission.
Until now, studies on the interaction between NoV and HBGAs have revealed that more than 10 of 33 genotypes bound to HBGAs, and wide recognition of HBGAs by GII/4 strains seems to be linked to transmission strength (see paragraph 3-5). On the other hand, no binding was observed in at least two NoV genotypes 10, 12, 17 and Shirato-Horikoshi, unpublished data. It is still unclear whether HBGAs act as the primary receptors or enhance NoV infectivity or attachment to a common cellular receptor. Further investigation is needed to clarify this point. Furthermore, sapoviruses, which are classified as members of the family Caliciviridae (Table 1) as noroviruses and are causative agents of human gastroenteritis, showed no binding to the HBGAs 19.



