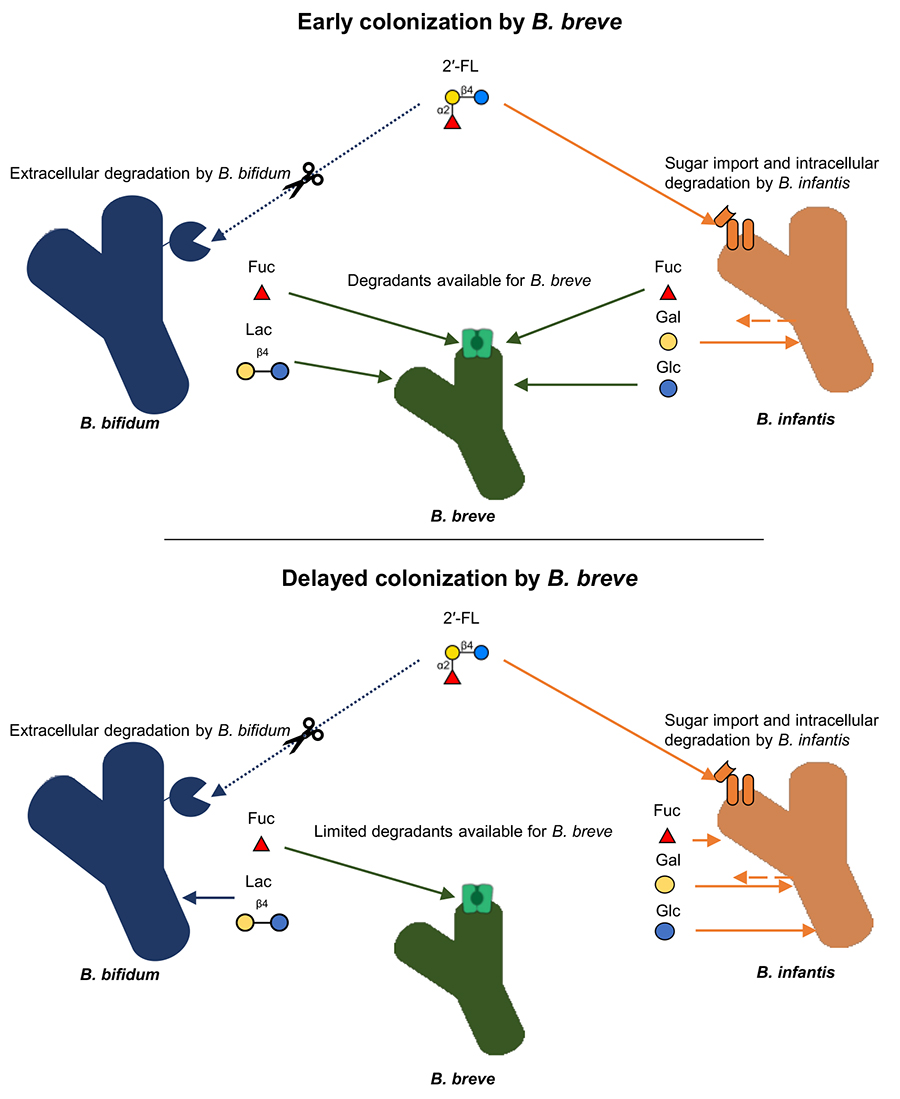
B. bifidum やB. infantisを最初に培地中に導入した場合は、予想通り、これらの種がビフィズス菌コミュニティーの優占種となった。後から移入してきた種が栄養源として利用可能なHMOs量が限られてしまうためである。しかしながら、 B. breve を先に培地中に導入した場合、予想に反して本菌種が最終的なコミュニティーにおける優占種となった。B. breve のHMOs利用能は限定されてるが、B. bifidum やB. infantisが資化過程で産生するHMOs構成糖(特にフコース)を利用することによってコミュニティーを優占したと考えられる。実際、B. bifidum とB. infantis のいずれもが、HMOsを分解利用する際にフコースを含む単糖を遊離することが知られている。B. bifidum は菌体外酵素で HMOs を分解するため、フコースとその他のHMOs分解物は細胞外で生じる36。B. infantis はHMOsをそのまま取り込み、フコースを含む単糖を一過的に細胞外へ放出することが報告されている37。そのため、早い段階でB. breveがそのニッチに存在していれば、B. breve は他の菌種がつくり出す分解物を利用することで優占種となり得るのである(図 2)。混合培養で観察されたこのような傾向は、in vivo のデータでも確認された。すなわち、ヨーロッパで行われた大規模な乳児コホートの糞便 DNA メタゲノムデータを解析したところ、出生直後にB. breve が検出されていた乳児では、4ヵ月後においてB. breve がビフィズス菌コミュニティーの優占種(50 %以上)となっている確率が有意に高かった38。これらの結果から、菌叢形成過程には先住効果が大きな影響を及ぼしていること、また、その過程はそれぞれの菌種が有する糖利用能によっても大きな影響を受けていることが明らかとなった。
図 2. B. breveがHMOs存在下で示す先住効果の予測図 2′-FL を例とした際のB. bifidum, B. infantis, およびB. breve による糖資化経路。その他のフコシル化HMOsも同様のメカニズムで資化されると推察される。B. bifidumの代謝経路は青、B. infantis ではオレンジ、B. breve では緑で示されている。菌による糖の取り込みは実線、酵素による糖の分解は点線、糖の放出は破線で示す。B. breve の多くの株は単独では2′-FLを資化することはできないが、早い段階でニッチに導入された場合、他の菌種がつくりだす2′-FL 分解物を利用してコミュニティーを優占することが出来る。一方で、導入が遅れると分解物は他の菌種によって消費されてしまうために優占することが出来ない。2′-FL, 2′-fucosyllactose; Lac, lactose; Fuc, Fucose; Glc, Glucose; GlcNAc, N-Acetylglucosamine; Gal, Galactose.
Ley, R. E. et al. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. 102, 11070–11075 (2005).
Kostic, A. D. et al. The dynamics of the human infant gut microbiome in development and in progression toward type 1 diabetes. Cell Host Microbe17, 260–273 (2015).
Qin, J. et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature490, 55–60 (2012).
Petersen, C. & Round, J. L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 16, 1024–1033 (2014).
Turroni, F. et al. The infant gut microbiome as a microbial organ influencing host well-being. Ital. J. Pediatr. 46, 1–13 (2020).
Underwood, M. A. et al. A comparison of two probiotic strains of bifidobacteria in premature infants. J. Pediatr. 163, 1585-1591.e9 (2013).
Plummer, E. L. et al. Gut microbiota of preterm infants supplemented with probiotics: sub-study of the ProPrems trial. BMC Microbiol. 18, 1–8 (2018).
Kitajima, H. et al. Early administration of Bifidobacterium breve to preterm infants: Randomised controlled trial. Arch. Dis. Child. Fetal Neonatal Ed. 76, 101–107 (1997).
Tissier, H. Recherches sur la flore intestinale des nourrissons (etat normal et pathologique). (University of Paris, Paris, France, 1900).
Tannock, G. W. et al. Comparison of the compositions of the stool microbiotas of infants fed goat milk formula, cow milk-based formula, or breast milk. Appl. Environ. Microbiol. 79, 3040–3048 (2013).
Matsuki, T. et al. A key genetic factor for fucosyllactose utilization affects infant gut microbiota development. Nat. Commun. 7, 11939 (2016).
Sakanaka, M. et al. Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria-infant symbiosis. Sci. Adv. 5, eaaw7696 (2019).
López, P., González-Rodríguez, I., Gueimonde, M., Margolles, A. & Suárez, A. Immune response to Bifidobacterium bifidum strains support Treg/Th17 plasticity. PLoS One6, (2011).
Kalliomäki, M. et al. Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing. J. Allergy Clin. Immunol. 107, 129–34 (2001).
Meng, D. et al. Indole-3-lactic acid, a metabolite of tryptophan, secreted by Bifidobacterium longum subspecies infantis is anti-inflammatory in the immature intestine. Pediatr. Res. 88, 209–217 (2020).
Verma, R., Lee, C. & Jeun, E. J. Cell Surface Polysaccharides of Bifidobacterium bifidum Induce the Generation of Foxp3 + Regulatory T Cells. Sci. Immunol. 103, 3–4 (2019).
Underwood, M. A. & Sohn, K. The Microbiota of the Extremely Preterm Infant. Clin. Perinatol. 44, 407–427 (2017).
Westerbeek, E. A. M. et al. The intestinal bacterial colonisation in preterm infants: A review of the literature. Clin. Nutr. 25, 361–368 (2006).
Magne, F., Suau, A., Pochart, P. & Desjeux, J. F. Fecal microbial community in preterm infants. J. Pediatr. Gastroenterol. Nutr. 41, 386–392 (2005).
Sakanaka, M. et al. Varied pathways of infant gut-associated Bifidobacterium to assimilate human milk oligosaccharides: prevalence of the gene set and its correlation with bifidobacteria-rich microbiota formation. Nutrients12, 71 (2020).
Ojima, M. N. et al. Ecological and molecular perspectives on responders and non-responders to probiotics and prebiotics. Curr. Opin. Biotechnol. 73, 108–120 (2022).
Ninonuevo, M. R. et all. A strategy for annotating the human milk glycome. J. Agric. Food Chem. 54, 7471–7480 (2006).
Engfer, M. B., Stahl, B., Finke, B., Sawatzki, G. & Daniel, H. Human milk oligosaccharides are resistant to enzymatic hydrolysis in the upper gastrointestinal tract. Am. J. Clin. Nutr. 71, 1589–1596 (2000).
Macrobal, A. & Sonnenburg, J. L. Human milk oligosaccharide consumption by intestinal microbiota. Clin Microbiol Infect18, 12–15 (2012).
Macrobal, A. et al. Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 58, 5334–5340 (2010).
Fukami, T. Historical contingency in community assembly: integrating niches, species pools, and priority effects. Annu. Rev. Ecol. Evol. Syst. 46, 1–23 (2015).
Chesson, P. Mechanisms of Maintaining Species Diversity. Annu. Rev. Ecol. Syst. 31, 343–66 (2000).
Costeloe, K., Hardy, P., Juszczak, E., Wilks, M. & Millar, M. R. Bifidobacterium breve BBG-001 in very preterm infants: A randomised controlled phase 3 trial. Lancet387, 649–660 (2016).
Satoh, Y. et al. Bifidobacteria prevents necrotizing enterocolitis and infection in preterm infants. Int. J. Probiotics Prebiotics2, 149–154 (2007).
Li, Y. et al. Effects of Bifidobacterium breve supplementation on intestinal flora of low birth weight infants. Pediatr. Int. 46, 509–515 (2004).
Horigome, A. et al. Colonization of Supplemented Bifidobacterium breve M-16V in Low Birth Weight Infants and Its Effects on Their Gut Microbiota Weeks Post-administration. Front. Microbiol. 12, 1–11 (2021).
O’Brien, C. E. et al. Early probiotic supplementation with B. infantis in breastfed infants leads to persistent colonization at 1 year. Pediatr. Res. 91, 627-636 (2022).
Debray, R. et al. Priority effects in microbiome assembly. Nat. Rev. Microbiol. 20, 109–121 (2022).
Sprockett, D., Fukami, T. & Relman, D. A. Role of priority effects in the early-life assembly of the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 15, 197–205 (2018).
Ojima, M. N. et al. Priority effects shape the structure of infant-type Bifidobacterium communities on human milk oligosaccharides. ISME J. 16, 2265-2279 (2022).
Gotoh, A. et al. Sharing of human milk oligosaccharides degradants within bifidobacterial communities in faecal cultures supplemented with Bifidobacterium bifidum. Sci. Rep. 8,13958 (2018).
Asakuma, S. et al. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J. Biol. Chem. 286, 34583–34592 (2011).
Bäckhed, F. et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe17, 690–703 (2015).