
氏名:中北 愼一
香川大学 医学部総合生命科学講座 准教授
1998年大阪大学大学院理学研究科博士課程修了後、大阪大学理学部有機生物科学講座(長谷純弘教授)に赴任、糖鎖構造解析及び糖鎖の蛍光標識法に関する研究に従事。2004年香川大学総合生命科学実験センター・糖鎖機能解析研究部門(香川県寄付講座)客員准教授、2008年香川大学総合生命科学実験センター・糖鎖機能解析研究部門・准教授、2020年香川大学医学部・総合生命科学講座・准教授、現在に至る。

氏名:平林 淳
東海国立大学機構名古屋大学 糖鎖生命コア研究所 特任教授
1982年東北大学修士取得後、帝京大学薬学部生物化学教室(笠井献一教授)に赴任、動物レクチンの研究に従事。1999年、GlycoXV(東京)にてグライコミクスを提起、2002年産業技術総合研究所(つくば)に異動、NEDO「糖鎖エンジニアリング」プロジェクト等に参画。2006年糖鎖医工学研究センター・副センター長、2012年幹細胞工学センター・首席研究員等を歴任。理学博士。現在、東海国立大学機構名古屋大学・糖鎖生命コア研究所にてプロジェクトマネージャー(Human Glycome)を担当。グライコームの着想(1998年12月)より四半世紀の年月を経て、ようやくその実現を迎えようとしている。日本糖鎖科学コンソーシアム 副会長、香川大学(2003~)、および横浜市立大学 客員教授(2019~)、香川県希少糖等共創推進会議-複合糖質・糖鎖部会 部会長(2024~)。
二糖類は、二つの単糖ユニットをもつ最も単純なオリゴ糖であり、理論的には、D 体および L 体のアルドヘキソースの組み合わせによって、最大 3,056 種類のユニークな構造を形成し得る。これらの大半はこれまで研究対象とされてこなかったが、近年の希少糖合成技術の進展により、実験的な探索が可能になりつつある(Nakakita SI, Hirabayashi J, BBA Advances, 7:100143, 2025)。この進展は、甘味料、医薬品、生分解性材料などへの応用が期待され、天然糖には見られない特性を有する二糖の発見につながる可能性を示している。より複雑なオリゴ糖と比較すると、二糖は体系的な研究が容易であり、構造と機能の関係を検証する上で適切な研究スケールに位置している。本稿では、新たにアクセス可能となったこの化学空間を探索するための体系的枠組みとして、「二糖グライコミクス」を提案し、科学的にも実用的にも重要な発見への道を拓くことを目指す。
二糖は、二つの単糖から構成されるオリゴ糖の中で最も単純な構造単位であり、さまざまな生物学的プロセスにおける基本的な認識単位として機能している。これまでの二糖研究は、主として自然界に豊富に存在する D-グルコース(D-Glc)、D-マンノース(D-Man)、D-ガラクトース(D-Gal)に由来するものにほぼ限定されていた。これに加えて、N-アセチル-D-グルコサミン(D-GlcNAc)、N-アセチル-D-ガラクトサミン(D-GalNAc)、L-フコース(L-Fuc)、D-キシロース(D-Xyl)、D-グルクロン酸(D-GlcA)といった誘導体も、天然糖鎖において一般的に見られる。重要な点は、これらの単糖およびその誘導体のほとんどが、D-Glc または D-Man を起点とする限られた生合成経路によって供給されていることである1,2。さらに、高等動物細胞に特徴的なシアル酸(9 炭素酸性糖)についても、その生合成は N-アセチル-D-マンノサミン(C6)とピルビン酸(C3)から成る、アルドール縮合に類似した機構によって進行する。
一方、理論的には、単糖レベルには自然界では利用されていない数多くの異性体、すなわちジアステレオマーが存在する。D-アルドヘキソースでは、D-アロース(D-All)、D-アルトロース(D-Alt)、D-タロース(D-Tal)、D-グロース(D-Gul)、D-イドース(D-Ido)がそれに該当する(図 1)。これらは一般に「希少糖」と呼ばれ、「自然界にほとんど存在しない単糖およびその誘導体」と定義されている3,4。また、L 系列に属するすべての単糖(アルドースおよびケトース)は、基本希少糖に分類される。この点において、糖鎖は核酸やタンパク質とは本質的に異なる。すなわち、糖鎖の構成要素である単糖は異性体として「数列的」に存在するのに対し、ヌクレオチドやアミノ酸には糖鎖のように数列的な多様性が存在しない。その結果、糖鎖に内在する膨大な立体化学的可能性の多くは未だ研究されてこなかった。しかし近年、酵素学および発酵科学の進展により、D-フルクトース(D-Fru)などの一般的な単糖から、L 系列を含む多様な希少糖へと変換する技術が利用可能になりつつある5-7。
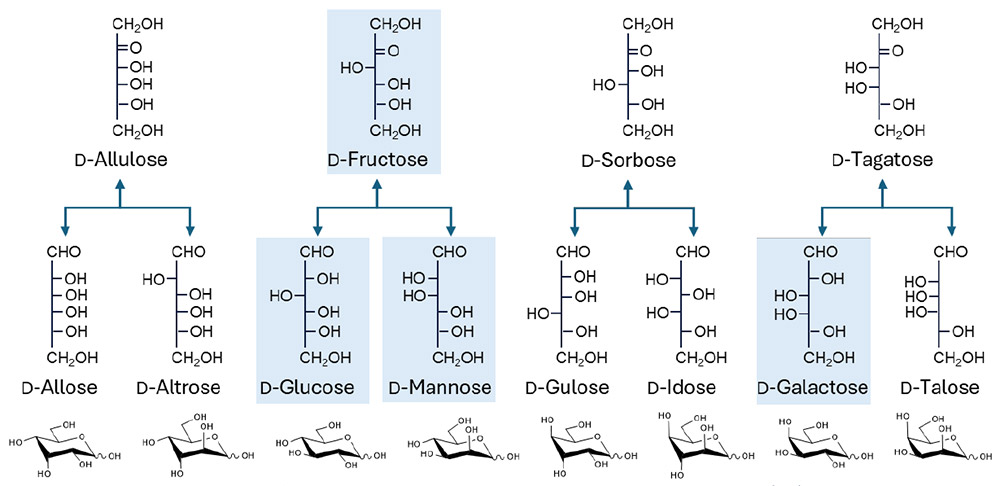
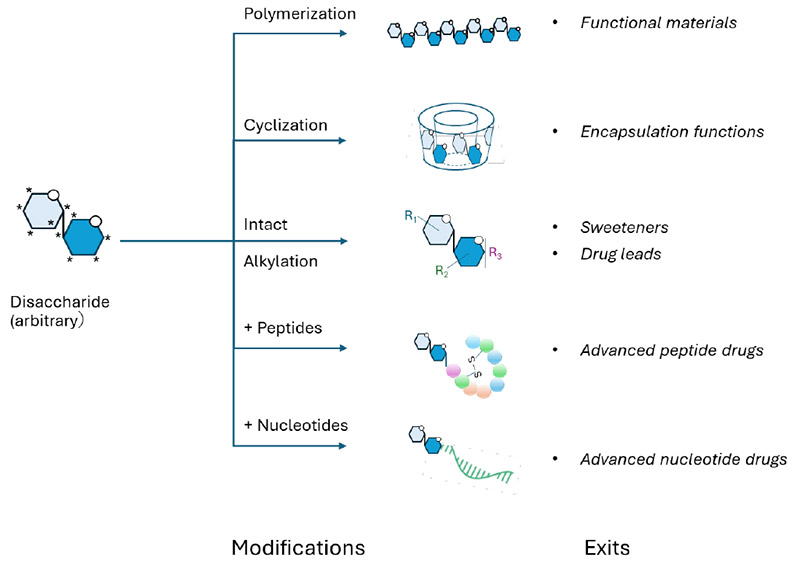
仮に、あらゆるアルドヘキソースがグリコシド結合を形成して二糖を構成できるとすると、その組み合わせ数は非常に大きくなる。中北と平林は、二つの未修飾アルドヘキソースから形成され得るユニークな二糖の総数、すなわち「二糖グライコームサイズ」が 3,056 であることを報告した注1)8。ここで、未修飾アルドヘキソースとは化学的に改変されていない単純な六炭糖を指し、「化学空間」は形成可能な分子構造の多様性範囲を表す注2)。糖鎖は単糖の連結様式が極めて多様であり、三糖になるだけで 10 万通りを超える組み合わせが生じる9。このことは、二糖レベルですら、その化学空間の多くが未開拓であることを示している。したがって、非天然型二糖の合成は、天然糖には見られない新規の物性や機能をもつ分子の発見につながる可能性を秘めており、医薬、バイオテクノロジー、材料科学など多様な分野への応用展開が期待される(図 2参照)。これに対して、三糖以上になると構造多様性は急激に増大し、そのすべてを包括的に取り扱うことは事実上不可能となる。これは、糖鎖研究における「組み合わせ爆発」として知られる現象であり、糖単位の数が増えるにつれて、可能な構造数が指数関数的に増大することを意味する。この観点から見ると、二糖グライコミクスは、複雑性が制御不能となる直前に位置する、実験的にアクセス可能な唯一の構造群を対象とした研究領域である。本研究では、化学的および酵素的な合成戦略を組み合わせ、多様な二糖を体系的に創出し、それらの性質を網羅的に解析する。このアプローチにより得られる知見は、糖鎖の基礎理解を深化させるとともに、新たな機能性分子の開発に向けた確かな基盤を提供するものである。
これらの分子に対して、融点や溶解性といった物理的特性に加え、「生体適合機能」、すなわち生物学的システムにおける適合性や役割について、包括的な解析が行われる。具体的には、タンパク質との相互作用能力や、既知の生体材料を模倣することによる細胞プロセスへの関与・介入能などが評価対象となる。こうして得られたデータは、オリゴ糖やさらに大きな糖鎖、ならびにそれらの複合体(glycoconjugates)の基本的性質に関する理解を深める上で重要な基盤となる。このような基礎的知見が蓄積されることで、二糖が持つ多様な応用可能性が系統的に検証され、実用化へとつながる道筋が明確化されていく。例えば、新規二糖の中には、標的型ドラッグデリバリーシステムの構築や、組織工学に資する革新的な生体材料の開発に利用されるものが現れる可能性があり、研究成果の社会実装に向けた展開が期待される。
注1)同じ二つの単糖が結合している場合、それらのα1-1βおよびβ1-1αの形は同一であるはずである。したがって、全てのアルドヘキソースの組み合わせの二糖類の総数は、16(非還元末端アルドヘキソース)× 12(結合様式;還元性二糖の場合はα1-2/3/4/6およびβ1/2/3/4/6、非還元性二糖の場合はα1-1α/α1-1β/β1-1α/β1-1β)× 16(還元末端アルドヘキソース)− 16 = 3,056、と計算される。
注2)化学空間には10の60乗を超える潜在的な分子が含まれており10、既知の化合物はごく一部に過ぎない。この広大な宇宙をナビゲートするために、研究者は分子量、脂溶性(log P)、極性表面積、水素結合能力などの記述子を座標として用いて構造をマッピングする11。この多次元的アプローチにより、新規医薬品候補を定量的に特定することが可能になる。
糖鎖はしばしば「生命の第三のコード」と呼ばれるが、そこに含まれる情報量は、核酸(DNA、RNA)やタンパク質といった線状高分子が担う情報をはるかに凌駕している。核酸やタンパク質とは異なり、糖鎖は高度に分岐した構造をとり得るため、その構造多様性と情報担持能は指数関数的に増大する。グライコミクスの観点から見ると、二糖は最小の情報単位、すなわち「ビット」と見なすことができる。実際には、二糖こそが、生物が分子認識やシグナル伝達において利用する「意味をもつ最小の構成単位」であると言える。
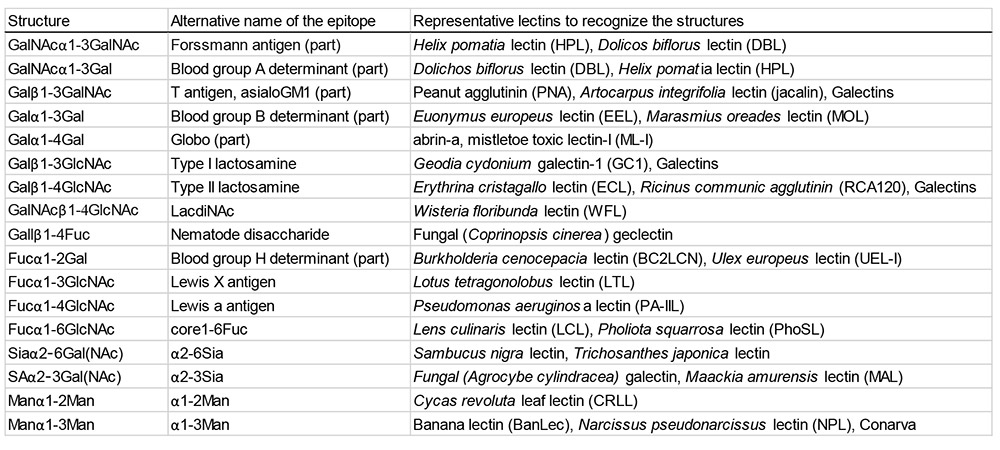
レクチンは、炭水化物(糖)に特異的に結合する遍在性タンパク質であり、細胞認識において不可欠な役割を担っている。レクチンはしばしば便宜的に単糖特異性に基づいて分類されるが12.13、実際には、ほとんどのレクチンが末端の単糖だけでなく、その一つ内側に位置する糖残基も同時に認識している。例えば、Ulex europaeus レクチンは血液型 H 抗原(Fucα1-2Gal)を認識し、これは血液型適合性において重要な意味をもつ。また、ジャカリンは T 抗原(Galβ1-3GalNAc)に結合する。このようなレクチン認識の特性を踏まえ、Albert M. Wu は糖鎖エピトープを二糖レベルで捉える分類概念を提唱し、体系的な命名法を導入するとともに、植物レクチンの特異性に基づいた二糖エピトープ研究を展開した14。同様に、ガレクチンに代表される動物レクチンは、ガラクトースと N-アセチルグルコサミンから成る二糖単位である N-アセチルラクトサミン(LacNAc)を基本的な認識モチーフとしている。現在では、「Galβ(syn)-gauche」と呼ばれる特定の立体配置を満たす二糖が、ガレクチンにより認識されるコンセンサス構造であることが明らかになっている2.15。代表的な二糖エピトープと、それを認識するレクチンの例を表 1に示す。
このように、生物学的な意味は二糖レベルで現れる。単糖が単なる「文字」として機能するのに対して、二糖は「単語」に相当する。文字が組み合わさることで言語として意味をもつ語が形成されるのと同様に、単糖から組み立てられた二糖は、生物学的情報を担う機能単位を形成する。新たに同定された二糖を体系的に特性評価することにより、糖鎖科学における比較の基盤が整備され、物性や機能の違いが生じる根本要因の理解や、機能予測へとつながる基礎情報が蓄積される。本研究は、このような新規の物理的特性および生物学的機能の発見機会を提供し、糖鎖研究におけるさらなる探究を、いっそう実りあるものにしていくだろう。
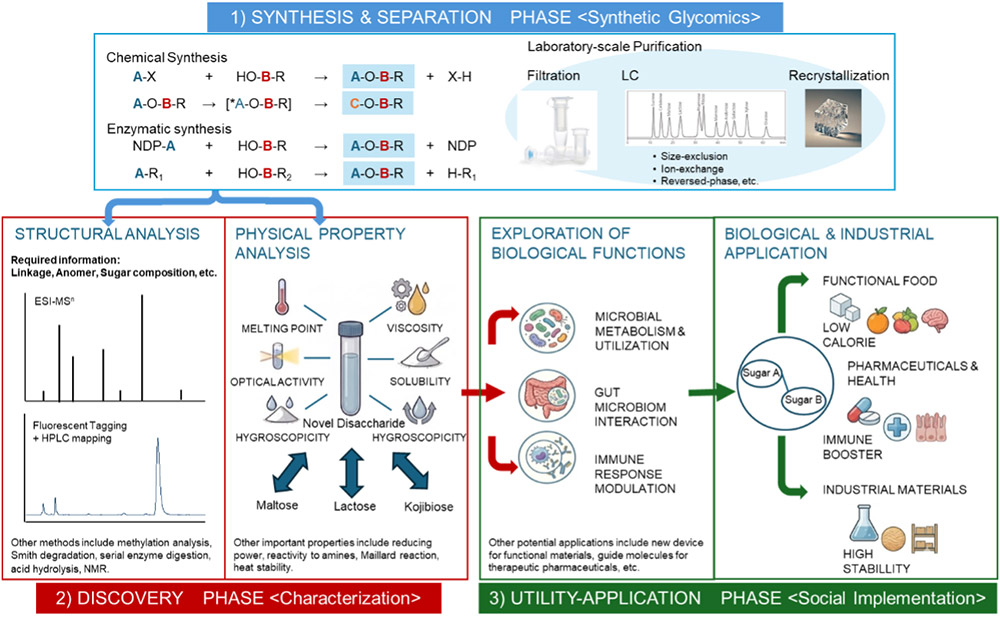
前述のとおり、本二糖グライコミクス・プロジェクトは、1)合成および分離、2)探索、3)利用応用という三つの主要な段階から構成されている(図 2)。それぞれの段階には固有の技術的課題が存在し、特に、酵素を用いた合成手法の確立や、高度な構造解析技術の整備は本分野の成否を左右する重要な要素である。これらの課題については、後述のコラム「非天然二糖類の酵素合成および構造解析における現状」において詳述する。
● 合成
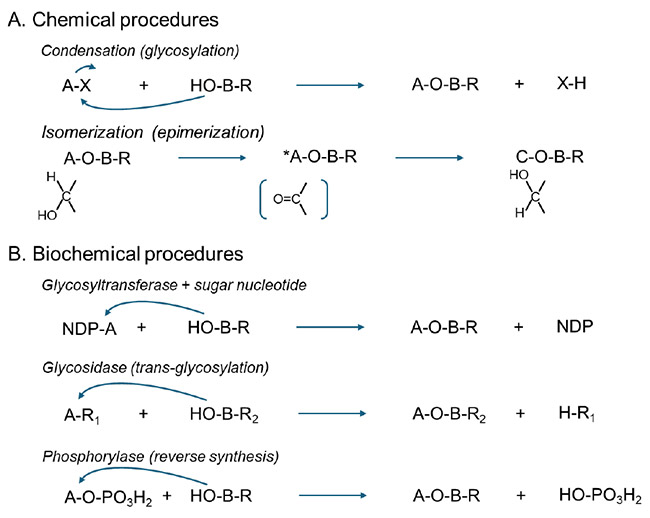
二糖は、化学的手法または酵素的手法のいずれによっても合成することができる(スキーム1)。化学合成法は一般に堅牢で再現性が高いが、3,000 種類以上に及ぶ新規二糖を網羅的に合成することは現実的ではない。化学合成によって得られる二糖標品は、分析法確立のための標準物質として、また物性解析や後続する探索段階における構造既知の参照物質として、特に有用であると考えられる。一方で、酵素を用いた二糖合成についても、これまでにいくつかの先行研究が報告されている¹⁶,¹⁷。しかしながら、酵素的アプローチは、希少糖にも作用する「特別な」酵素―すなわち、糖転移酵素、グリコシダーゼ、あるいはホスホリラーゼ―の探索に大きく依存しており、二糖を体系的かつ効率的に合成するための一般的手法を確立することは容易ではない。
● 分離
得られた二糖は、その構造同定および物理化学的特性評価を行うため、十分な純度にまで分離・精製される必要がある。この目的のため、サイズ排除、イオン交換、逆相、親水性相互作用クロマトグラフィーなど、多様な液体クロマトグラフィー手法の適用が検討される。分離に際しては、二糖を未修飾のまま用いるか、適切な試薬で標識した形で解析するかの選択が重要となる。二糖グライコミクスにおいては、効果的かつ汎用的な分離技術の確立がとりわけ重要である。特に希少糖は化学的安定性が低い場合が多く、還元末端に希少糖を含む二糖では、事前にグリコシド化するなどの配慮が不可欠となる。先行研究では、グルコース二糖類の反応性が結合様式(αまたはβ、ならびに 1-2、1-3、1-4、1-6)に大きく依存することが示されているが、その詳細な要因はいまだ解明されていない18。
● 構造解析
二糖を正確に特性評価するためには、単糖組成、アノメリック配置(α または β)、ならびにグリコシド結合位置を厳密に決定する必要がある。構造解析においては、NMR 分光法が最終的な構造決定において中心的役割を果たす。一方で、蛍光標識と高性能液体クロマトグラフィー(HPLC)を組み合わせることにより、異性体二糖の効率的な分離が可能となる。これに加え、MS/MS や MSⁿ 解析を含む質量分析(MS)手法、ならびにイオンモビリティ分離などの新規分離モードを活用することで、二糖の特性に関する新たな知見が得られる可能性がある19。さらに、結晶学的解析は、二糖がもつ固有の構造的特徴を原子レベルで明らかにする情報を提供する20。これらの手法を相補的に用いることにより、従来法の限界を克服し、体系的かつ信頼性の高い二糖解析が可能となり、グライコミクス研究の進展に寄与する。
● 物理特性の解析
多くの合成二糖は新規化合物であり、その物理特性に関する既存データは限られているが、新たな特性が見いだされれば、産業利用への展開可能性が大きく広がる。我々は、融点・沸点、溶解性、揮発性、吸湿性、旋光度、結晶性、甘味といった官能的特性を含む基本的な物理特性について、体系的な比較解析を行う予定である。このような包括的な物性プロファイリングにより、化学構造と物理化学的挙動との相関関係が明らかとなり、やがては、より広範な支配原理の解明へとつながることが期待される。
● 生体適合性機能の探索
新規二糖については、ラクトース(Galβ1-4Glc)、マルトース(Glcα1-4Glc)、セルロビオース(Glcβ1-4Glc)など既存の代表的二糖と比較しながら、微生物による認識や代謝、腸内細菌叢への影響、さらには免疫応答への作用を評価する21。前項で解析する物理的基本特性――融点、粘度、光学活性、溶解性、吸湿性、還元性、アミン反応性、熱力学的安定性、結晶化挙動、甘味――は、生物学的特性とも相関する可能性がある。本解析では、微生物による代謝および資化性、ならびにそれがもたらす生理学的影響に着目し、物理特性との関係性や潜在的な機能、さらには商業利用の可能性を探索する。近年では、GalNAc 修飾 siRNA が優れた標的指向性を示すなど、この分野における応用例も着実に現れつつある22。
● 価値のある二糖類の大規模生産(産業への橋渡し)
新規二糖の機能開発と並行して、D-グルコースやショ糖といった、安価で天然に豊富に存在する原料から価値のある二糖を生産可能な微生物(あるいは酵素系)の開発を目指す。具体的には、新規二糖を認識し、加水分解あるいは転移反応を触媒するグリコシダーゼや、それに関連する糖ヌクレオチドトランスポーター、グリコシルトランスフェラーゼ、レクチンなどを想定している。これらの生体触媒系の確立は、研究成果を産業応用へと橋渡しする上で重要な役割を果たす。
近年の酵素工学および発酵技術におけるブレークスルーにより、希少糖の生産が現実的なものとなった3-8。この動きは、生体内に存在する糖分子のみならず、それを超えた全糖分子の集合を解析対象とする「拡張グライコミクス」という概念と密接に関係している23。研究者はこれにより、理論上存在し得る 3,056 種類すべての二糖について、その性質と潜在的用途を体系的に評価することが初めて可能となったのだ。これは、従来の糖鎖研究と比較して、研究対象領域を飛躍的に拡張するものである。本研究のアプローチは、アルドヘキソースのすべての組み合わせに限定されるものの、それらの大部分は人工的に合成される糖であり、この点において新たに「合成グライコミクス」と呼ぶべき研究領域の成立を宣言するものである。合成グライコミクスは、人工糖の体系的創出と解析を通じて、創薬、高度バイオマテリアル、診断技術における革新への道を切り拓く可能性を有している。
ここで最も重要な概念の一つが、「糖鎖の化学空間」である。糖鎖の化学空間は、タンパク質や核酸のそれと比べても桁違いに大きい9。この圧倒的な多様性は、各単糖分子に多数存在するヒドロキシ基(–OH)と、それらが結合し得る多様な様式—結合位置、立体配置、アノメリック形態—に由来する。こうした特徴は、炭水化物を表す一般式(C·H2O)にすでに内在するもので、多数の水酸基と不斉炭素に富んだ構造に起因している1,2。二糖であっても、その化学空間はすでに爆発的なのだ。n 種類の単糖からなる場合、理論上の二糖数は n × n ×(結合位置)×(α/β)×(立体化学)となり、数千から数万種の潜在構造が生じる。これに対し、アミノ酸二量体は 1,600 通り(40 × 40)、核酸二量体は 16 通り(4 × 4)に過ぎず、糖鎖の多様性がいかに際立っているかが分かる。中北と平林は、アルドヘキソース(6個の炭素原子とアルデヒド基を持つ単糖)に限定した場合でも 3,056 種類の二糖が存在し得ることを示した8。なお、この数にはペントース、ケトース、さらには N-アセチル化体や O-硫酸化体などの派生構造は含まれておらず、これらを考慮すれば化学空間はさらに拡大する。
このように、二糖は糖鎖の化学空間における最初の「臨界点」を形成している。三糖になると構造多様性は急激に増大し、10 万種から 100 万種を超える可能性が生じる。実際、わずか 6 種類の一般的な単糖が連結するだけでも、その構造組み合わせは最大で 1.05 × 1012 通りに達するとの試算がある24。このため、二糖は人工糖の理解を深め、合成グライコミクスの概念を発展させるための最適な出発点と位置づけることができる。より小さく、扱いやすい構造単位から研究を開始することで、希少糖を含む合成糖の可能性を体系的に引き出し、科学および技術の両面において新たな発見へとつながる道筋が拓かれる。
二糖グライコミクスは、単糖から二糖への糖変換を体系的に推進することを目的としている。本分野における主要な課題は、二糖の合成および分離技術の確立に加え、新規二糖が示す機能の同定にあり、これらを克服するには高度な技術力と十分な研究資金が不可欠である。さらに、新規糖鎖製品の有用性は、実験的検証を経るまでは明らかにならないため、不確実性を内包している点も無視できない。糖質は、その構造的複雑さゆえに、タンパク質や核酸と比較して理解が著しく遅れており、本領域には未解明の課題が数多く残されている。例えば、糖質の起源や糖鎖が担う生物学的機能に関する理解は、依然として発展途上にある。一方で、国内においては、「ヒトグライコームプロジェクト」が、「拡張セントラルドグマ」を科学的目標として掲げている点が注目される25。二糖は、今後のグライコミクス研究における基盤的アプローチとして、極めて重要な役割を担うことになるだろう。
糖鎖はしばしば、その分岐構造から樹木にたとえられるが、二糖単位にまで分解することで初めて体系的理解が可能となる。二糖は、十分な構造多様性と情報量を備えた最小の機能単位である。すなわち、単糖では情報量が不足し、より大きな糖鎖では解析の複雑性が急激に増大する。情報理論の観点から見ると、二糖は「圧縮可能な単位」として最適であり、合成、分離、質量分析、ライブラリ構築、生物学的アッセイといった実験技術の射程内で扱うことができるサイズに位置している。また、二糖は化学空間における多様性が急激に拡大する閾値にあり、生物学的認識においても最小単位として機能する。このような特性から、二糖は糖鎖科学における主要な研究対象となるだけでなく、ネットワークの数学的モデリングやグラフ理論に基づく解析にも適した対象と考えられる。
構造的多様性に加え、二糖のもう一つの重要な利点は、コンパクトで高い親水性をもつ分子モジュールとしての性質にある。分子量は約 400 Da 程度と比較的小さいにもかかわらず、二糖には多数のキラル中心と高密度に配置されたヒドロキシ基が含まれている。この特性により、二糖は複数の機能成分を効率的に結合するリンカー、あるいはハブ分子として機能し得る。特に、立体化学的多様性、水溶性、ならびに精密な配座制御が求められる糖修飾オリゴヌクレオチドやペプチド治療薬など、近年注目される中分子創薬モダリティの発展において、二糖は重要な役割を果たす可能性がある。この意味において、二糖は将来の糖統合型中分子医薬設計に向けた「特権的足場(privileged scaffold)」を構成する分子基盤になるのではないか(図 3)。
二糖グライコミクスは、近年の希少糖合成技術の進展を背景として発展しつつある新たな糖鎖科学分野であり、二糖の構造、物理化学的特性、生物学的機能を体系的に解析することを目的としている。本分野は、精密な構造解析、機能評価、ならびに微生物学的アプローチを統合することにより、食品科学、医薬・製薬研究など多岐にわたる応用展開が期待される。グライコミクスおよびグライコインフォマティクスと密接に連携することで、二糖の包括的解析は糖鎖生物学の理解を一層深化させるとともに、新規機能性材料の創出を促進する基盤となる。グライコミクスの概念が提唱されてから 25 年以上が経過した現在においても、本領域は進化と拡張を続けている26。二糖グライコミクスはその次なる展開を切り拓く重要な位置を占めるものと考えられる。
本レビュー執筆における貴重な洞察と指導に対し、笠井献一博士に心より感謝申し上げます。また、モルテン・タイセン=アンデルセン博士(マッコーリー大学、名古屋大学)、ヤン・ゲラルデル博士(CNRS、岐阜大学)、水野真盛博士(野口研究所)、岩城隼博士(東京化成工業株式会社)、高津吉広博士(生化学工業株式会社)、西風隆司博士(株式会社島津製作所)、徳田雅明博士(香川大学)にも、二糖類グライコミクスへの有益なコメントと温かい支援に感謝いたします。
希少糖を含有する二糖類は、糖転移酵素(GTase)、トランスグリコシル化活性を有するグリコシダーゼ、あるいはホスホリラーゼといった酵素を利用することで合成可能である。このような酵素的手法は、化学的には合成が困難な非天然型二糖類の位置選択的および立体選択的構築を実現し、合成グライコミクス分野の発展に寄与している。
i. グリコシルトランスフェラーゼ(GTase)
GTaseはアノメリック構造および結合位置に対する高い制御能力を有しており、多くの細菌由来GTaseは大腸菌において組換え発現が可能である1-5。しかしながら、糖ヌクレオチド供与体の調製コストの高さや基質特異性の厳格さが、その応用範囲を著しく制限している。
ii. グリコシダーゼによるトランスグリコシル化
トランスグリコシル化は、p-ニトロフェニル単糖(pNP糖)をドナー基質として利用することで、理論的には単一の反応プロセスによって多種多様な二糖異性体の合成が可能となる多機能的アプローチとみなされている6。一方、非還元性三糖の生成については想定外の事象として報告されている7。現在、新規グリコシダーゼによるトランスグリコシル化活性の体系的探索が進行中である8が、主な課題として、希少糖に対する触媒活性酵素の不足、希少糖pNPドナーの限定的供給、ならびに標準化された精製手法の未整備が指摘される。希少糖の化学的安定性が低いことから、効率的で温和な分離プロセスの開発が必須となる。
iii. ホスホリラーゼ基盤の合成
ホスホリラーゼは、可逆的なホスホリシスおよび縮合反応を触媒する酵素であり、ラクト-N-ビオースの事例にみられるように大規模生産プロセスへの応用が可能とされる9-11。しかしながら、既知のホスホリラーゼの多くはグルコース特異性を有しており、希少糖受容体の非還元末端に対するグルコシル化反応には利用範囲が限定されるという課題が存在する。
質量分析に基づく手法(MS/MS または MSn)では、ラベルなしまたは適切にラベルされた二糖類に対して、断片化パターンを詳細に系統的に解析する必要がある。また、イオンモビリティ法など、新しい質量分離モードを使用することで二糖異性体相互の分離と識別が可能になるかもしれない。さらに、NMR 分光法は溶液中で包括的で非破壊的な構造情報を提供し、X 線結晶解析は原子レベルでの分子間相互作用を明らかにする。



