

|
|
糖鎖遺伝子の分子進化 |
|||||||||||
 |
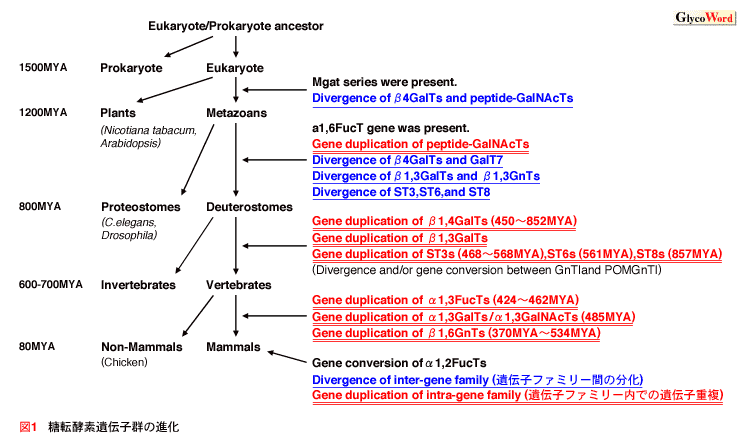
20世紀の終盤から、各動物種のゲノム計画の発展の恩恵を受け、糖鎖遺伝子がデータベースを元に次々とクローニングされた。これまでに160種類を越える糖転移酵素がクローニングされ、今もまだその数を増やしており、近い将来には200種類を越えると言われている(Narimatsu, Glycoconj. J. 2004)。糖転移酵素は糖を糖ヌクレオチド(donor、糖供与体)から伸長される糖鎖(acceptor、受容体)へ付加する反応を触媒する。転移酵素のアミノ酸配列は、糖供与体、受容体、結合様式などの酵素の特異性を反映する。従って、糖転移酵素は、その塩基配列の相同性に基づいて機能的な遺伝子ファミリーに分類することができる。様々な糖転移酵素遺伝子ファミリーの解析結果から、触媒する結合様式は厳密で、糖供与体に対しての基質特異性は比較的高く、受容体に対しての基質特異性にはかなり柔軟性があることが分かってきた。 本稿では個々の糖転移酵素遺伝子ファミリーに着目して遺伝子の進化を論じるのではなく、糖転移酵素遺伝子全体の進化の傾向について論じたい。我々はN-結合型、O-結合型糖鎖合成に関与する代表的な糖転移酵素遺伝子についてアミノ酸配列用いて系統樹解析を行い、各糖転移酵素遺伝子の分岐年代を推定した(Kaneko, TIGG, 2000)。その結果、遺伝子ファミリー内の遺伝子重複が脊椎動物の分岐後に集中していることが明らかとなった。また、幾つかの遺伝子ファミリー間では、この遺伝子ファミリー内の遺伝子重複が起こる前に、遺伝子ファミリー間での遺伝子重複が起こったことが示唆された(図1)。つまり、まだ遺伝子の数が少ない祖先型遺伝子の段階で遺伝子重複を起こし、機能を分化させ、異なる活性を持つ別々の祖先遺伝子となり、その後、それぞれがまた遺伝子重複をしてその数を増やし、現在の形となったと考えられる。 |
||||||||||
 |
|||||||||||
|
ところで、脊椎動物出現時期と顎口類出現時期の2回にゲノム重複が起こったことが示されている(Kasahara, Trend in Genetics,
1997)。我々は、糖転移酵素遺伝子群が「ホモロガスクラスター」を形成している痕跡を見出した。「ホモロガスクラスター」とは、ゲノムに2カ所以上散在した相同領域の事で、2回のゲノム重複で4倍体化したゲノム進化の痕跡である。例えば、6番染色体のMHC領域に相同な領域が染色体1、9、19番上に見出される。また、Hoxクラスターは2,7,12,17番染色体上に見出される。α1,3フコース転移酵素遺伝子は、6,9、19番染色体に5つの遺伝子ファミリーが散在している。また、β1,3ガラクトース転移酵素遺伝子群は1,6,19番染色体に4つの遺伝子が存在する。Mgat遺伝子群は染色体2,7,12,17番に存在し、シアル酸転移酵素遺伝子群は1,9,2,12,17番染色体上に見出される。これら糖転移酵素遺伝子の「ホモロガスクラスター」の存在は、先程述べた系統樹解析によって得られた遺伝子重複の時期と矛盾しない。つまり、糖転移酵素遺伝子のおおよそ基本的なセットはゲノム重複を起こす前のゲノムに揃っており、その後、2回のゲノム重複により、一気にその遺伝子の数を増やし、各遺伝子ファミリー内においての酵素活性の多様性を獲得したと思われる。この染色体の位置情報は、周辺の遺伝子との位置関係などと併せて、遺伝子配列だけからは推定出来ない、より長い時間の糖転移酵素遺伝子の進化を考察する助けとなるだろう。 |
|||||||||||
|
<追記>各種糖転移酵素遺伝子が取得された際、しばしば、系統樹を作成し、新規の糖転移酵素遺伝子と他の糖転移酵素遺伝子の関係が論じられる。そのような系統樹作成の為の分子進化学的解析を行ったことのある人ならば、比較に使用する遺伝子(OTU)の本数を変えたり、比較する領域を変えたりした場合、遺伝子間の相対的位置関係が変わってしまうような現象をみたことがあると思う。また、時々、とても遠い関係の遺伝子群を比較して、系統樹を示している場合も見受けられる。果たしてそのような不確かな解析で得られた系統樹に意味があるのだろうか?分子進化の系統樹作成のソフトは、単に文字列を比較して距離を計算しているだけなので、意味のない比較でも計算は出来るし、系統樹を描かせることも出来る。単なる文字情報としての「配列」を安易に比較することのないよう、注意を喚起したい。 |
|||||||||||
| 金子美華(山形大学医学部 環境病態統御学講座・法医病態診断学分野) 斎藤成也(国立遺伝学研究所 集団遺伝研究部門) |
|||||||||||
|
|||||||||||
|
|
|||||||||||
| 2005年3月14日 | |||||||||||
|
|
|||||||||||
|
|||||||||||
