

|
|
歁擕椶偺庴惛偲摐嵔 |  | |||||||||||||
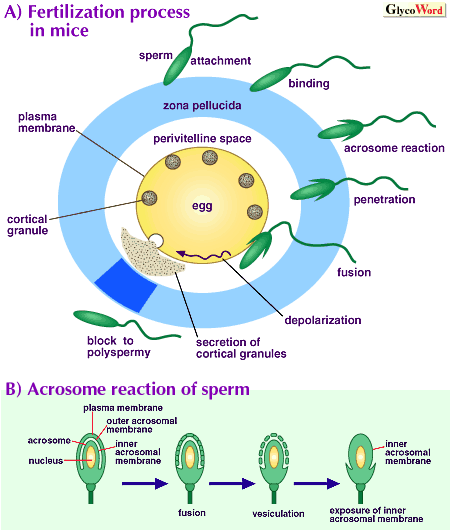
 | 丂歁擕椶偺庴惛夁掱偵偼丄條乆側惛巕偲棏偺憡屳嶌梡偑娷傑傟偰偄傞乮恾乯丅庴惛擻傪妉摼偟偨惛巕偼丄偼偠傔棏傪旐偭偰偄傞摟柧懷偵庛偔晅拝偟丄傗偑偰傛傝嫮偔寢崌偡傞傛偆偵側傞丅惛巕乕棏偺寢崌偼丄棏摟柧懷偺摐僞儞僷僋幙偲偙傟偵懳偡傞惛巕枌昞柺偺寢崌僞儞僷僋幙偲偺摿堎揑側憡屳嶌梡傪夘偟偰峴側傢傟傞丅棏摟柧懷傊偺寢崌偼丄惛巕偺愭懱奜枌偲嵶朎枌偺梈崌丄枌彫朎偺宍惉偲偦傟傜偺扙棊傪宱偰愭懱撪枌偺業弌乮愭懱斀墳乯傪桿摫偡傞丅愭懱斀墳桿摫惛巕偼丄摟柧懷傪娧捠偟偰埻棏峯偵払偟丄棏嵶朎枌偲梈崌偡傞丅梈崌偟偨棏嵶朎偵偼丄懡惛傪慾巭偡傞斀墳偲偟偰丄媫懍側枌偺扙暘嬌偲丄棏嵶朎撪偺儕僜僝乕儉條峺慺偵晉傓昞憌梓棻偺埻棏峯傊偺暘斿偑桿摫偝傟傞丅偦偺寢壥丄摟柧懷偺暔棟揑丄壔妛揑曄壔偲惛巕寢崌妶惈偺憆幐偑傕偨傜偝傟傞丅 | |||||||||||||
 | ||||||||||||||
|
丂偙偺傛偆偵丄棏摟柧懷偼惛巕偲偺寢崌丄惛巕愭懱斀墳偺桿摫丄懡惛慾巭偵偍偄偰廳梫側栶妱傪扴偭偰偄傞偑丄偦偺慻惉偼妱崌扨弮偱偁傞丅儅僂僗偱偼ZP1丄ZP2丄ZP3偲屇偽傟傞俁庬偺摐僞儞僷僋幙丄僽僞偱偼90K丄55K兛丄55K兝偲屇偽傟傞俁庬偺摐僞儞僷僋幙偐傜峔惉偝傟偰偄傞丅僸僩丄僒儖丄僴儉僗僞乕丄儔僢僩丄僂僒僊摍偵偍偄偰傕尷傜傟偨庬椶乮俀乕俁庬乯偺摐僞儞僷僋幙偑娷傑傟偰偄傞偵夁偓側偄丅儅僂僗偺宯偱偼丄惛巕偲偺寢崌傗惛巕愭懱斀墳偺桿摫偵偼ZP3偑丄愭懱斀墳傪桿摫偟偨惛巕偲偺寢崌偵偼ZP2偑娭梌偡傞偙偲偑帵偝傟偰偍傝丄傑偨僽僞偱偼55K兛偲55K兝偺暋崌懱偑惛巕偲偺傛傝嫮偄寢崌偵娭梌偡傞偙偲偑帵嵈偝傟偰偄傞丅棏摟柧懷摐僞儞僷僋幙偺摐嵔峔憿偵娭偟偰偼丄僽僞偺俶寢崌宆偍傛傃俷寢崌宆摐嵔丄儅僂僗丄僂僔偺俶寢崌宆摐嵔偺忣曬偑摼傜傟偰偄傞丅 丂庴惛偼婎杮揑偵偼庬摿堎揑偵峴側傢傟傞丅椺偊偽丄僸僩偺惛巕偼儅僂僗傗僴儉僗僞乕偺棏偵偼寢崌偟側偄丅偙傟偼棏摟柧懷摐僞儞僷僋幙忋偺摐嵔傪擣幆偟偰寢崌偡傞応崌偺摿堎惈偺堘偄偵傛傞傕偺偲峫偊傜傟偰偄傞丅嵟嬤丄敪惗岺妛揑曽朄偵傛偭偰儅僂僗ZP3偺戙傢傝偵僸僩偺ZP3傪敪尰偝偣偨儅僂僗偑嶌惢偝傟丄偙偺棏偵偼僸僩偺惛巕偼寢崌偱偒側偄偑丄儅僂僗偺惛巕偼寢崌偱偒傞偙偲偑帵偝傟丄庬摿堎揑側庴惛偵偍偗傞棏摟柧懷摐僞儞僷僋幙偺摐嵔峔憿偺廳梫惈偑夵傔偰巜揈偝傟偰偄傞丅惛巕庴梕妶惈傪扴偆摐嵔偵娭偟偰偼丄庡偵儅僂僗偺宯傪拞怱偵尋媶偑恑傔傜傟丄俀偮偺愢偑採彞偝傟偰偒偨丅戞堦偼丄兛-Gal巆婎傪娷傓俷寢崌宆摐嵔傪擣幆偡傞斀墳偺夘嵼傪帵偡Wassarman愢偱丄偙偺傛偆側摐嵔偲寢崌偡傞僞儞僷僋媛偲偟偰惛巕懁偺sp56偑帵嵈偝傟偰偒偨丅偟偐偟丄兛-僈儔僋僩乕僗揮堏峺慺寚懝儅僂僗偑庴惛擻傪桳偡傞偙偲丄sp56偑惛巕偺嵶朎昞柺偵偼業弌偟偰偄側偄偙偲摍丄偙偺峫偊偵柕弬偡傞帠幚傕曬崘偝傟偰偄傞丅戞擇偼丄惛巕昞柺偺兝-僈儔僋僩乕僗揮堏峺慺偑棏懁偺 GlcNAc 傪旕娨尦枛抂偵帩偮俷寢崌宆摐嵔傪擣幆偟偰寢崌偡傞偲偄偆Shur 愢偱偁傞丅嵟嬤丄兝-Gal揮堏峺慺 I寚懝儅僂僗偑嶌惢偝傟偨偑丄偙偺儅僂僗偺惛巕偼 in vitro偱偺ZP3傊偺寢崌擻偑栰惗宆偺30-50%偵尭彮偟丄傑偨ZP3偵傛傞愭懱斀墳偺桿摫偼寚擛偟偰偄傞偵傕偐偐傢傜偢丄 in vivo偱偼惓忢側庴惛擻傪桳偡傞偲偄偆丅傑偨丄忋婰偺愢偲偼堎側傝丄棏懁摐嵔偺兝-Gal巆婎傪擣幆偡傞斀墳偺廳梫惈傪帵偡曬崘傕偁傝丄嵟傕尋媶偝傟偰偄傞儅僂僗偺宯偱偡傜惛巕乕棏娫偺摐嵔擣幆婡峔偼埶慠偲偟偰柧妋偵偼側偭偰偄側偄丅 丂庴惛偵偍偄偰摐嵔偑嬌傔偰廳梫側栶妱傪墘偠偰偄傞偙偲偵媈偄偼側偄偑丄惛巕庴梕懱摐嵔偺峔憿偺摨掕傗摿挜晅偗丄惛巕懁偺摐嵔擣幆暘巕偺摨掕摍丄懡偔偺栤戣偑嶳愊偟偰偄傞丅偦傟傜偺夝柧偼丄庴惛偺庬摿堎惈傗懡惛慾巭婡峔傪柧偐偵偡傞忋偱丄嬌傔偰廳梫側壽戣偲側偭偰偄傞丅 | ||||||||||||||
| 崅嶈丂惤堦乮搶嫗戝妛堛壢妛尋媶強丒嵶朎惗暔壔妛尋媶晹乯 | ||||||||||||||
| ||||||||||||||
| 1998擭 12寧 15擔 | ||||||||||||||
| ||||||||||||||

