

|
|
兝-1,4-僈儔僋僩乕僗揮堏峺慺僼傽儈儕乕 | |||||||||||||||||
 |
丂惗暔妶惈傪桳偡傞摐嵔峈尨偺懡偔偼丄摐嵔懁嵔偵尒傜傟傞Gal兝1-->3/4GlcNAc偲偄偆婎杮峔憿偵敪尰偟偰偄傞丅兝-僈儔僋僩乕僗揮堏峺慺(兝-GalT)偼丄UDP-Gal偐傜Gal傪摐嵔枛抂偺GlcNAc巆婎偵揮堏偡傞偙偲偱偙偺擇摐峔憿傪嶌傝弌偡偑丄嵟嬤偵側偭偰偙傟傑偱偵抦傜傟偰偄側偄兝-1,4-GalT傗兝-1,3-GalT傪僐乕僪偡傞堚揱巕偑師乆偵僋儘乕僯儞僌偝傟丄偙傟傜傕偦傟偧傟僼傽儈儕乕傪宍惉偟偰偄傞偙偲偑柧傜偐偵側偭偰偒偨丅偟偐偟丄兝-1,4-GalT偲兝-1,3-GalT偼丄婎幙偲偟偰摐嫙梌懱傗摐庴梕懱傪嫟桳偡傞偵傕偐偐傢傜偢丄偙傟傜偺摐揮堏峺慺娫偱偼憡摨惈椞堟偼尒弌偝傟偰偄側偄丅杮峞偱偼丄惗懱撪偵偦偺峺慺妶惈偑峀偔暘晍偡傞兝-1,4-GalT僼傽儈儕乕偵徟揰傪偁偰夝愢偡傞丅 丂兝-1,4-GalT堚揱巕偼丄1986擭偵僂僔恡憻偲擕態偺cDNA儔僀僽儔儕乕偐傜僋儘乕僯儞僌偝傟偨(兝-1,4-GalT I)丅偦偺屻丄偙偺堚揱巕偺儂儌儘乕僌偑扨棧偝傟側偐偭偨偙偲偐傜丄兝-1,4-GalT偼惗懱撪偵偼堦偮偟偐懚嵼偣偢丄偦傟偑摐僞儞僷僋幙摐嵔偵尒傜傟傞慡偰偺兝-1,4-寢崌偟偨僈儔僋僩乕僗偺揮堏傪扴偭偰偄傞偲峫偊傜傟偰偒偨丅偲偙傠偑丄兝-1,4-GalT I堚揱巕攋夡儅僂僗偺奺慻怐傗寣惔偺摐僞儞僷僋幙偵偼丄摐嵔枛抂偵兝-1,4-寢崌偟偨Gal巆婎偑尒弌偝傟丄傑偨慻怐儂儌僕僃僱乕僩拞偵兝-1,4-GalT妶惈偑巆懚偡傞偙偲側偳偐傜丄惗懱偵偼暿偺兝-1,4-GalT偑懚嵼偡傞偙偲偑帵嵈偝傟偨丅 丂怴婯偺僸僩兝-1,4-GalT堚揱巕偼丄摐揮堏峺慺娫偵尒傜傟傞傾儈僲巁攝楍偺儂儌儘僕乕(1)傗堚揱巕僶儞僋偵搊榐偝傟偨堚揱巕抐曅偺攝楍忣曬(2, 3)傪棙梡偟偰僋儘乕僯儞僌偝傟偨丅偦傟傜偼兝-1,4-GalT I偲傾儈僲巁攝楍偱55, 44, 41, 37, 33%偺儂儌儘僕乕傪帩偭偰偄偨偺偱丄杮峞偱偼儂儌儘僕乕偺弴斣偵廬偭偰兝-1,4-GalT II, III, IV, V, VI偲婰嵹偡傞丅偟偐偟丄杮峞偺兝-1,4-GalT V偼暥專(1)偺兝-1,4-GalT IV偵憡摉偡傞偺偱拲堄偝傟偨偄丅僸僩兝-1,4-GalT僼傽儈儕乕偺偆偪丄兝-1,4-GalT I偼儅僂僗偲僯儚僩儕丄兝-1,4-GalT II偼僯儚僩儕偐傜傕偦偺cDNA偑扨棧偝傟偰偄傞丅偛偔嵟嬤丄儔僋僩僔儖僙儔儈僪崌惉峺慺偑儔僢僩擼偐傜惛惢偝傟丄偦偺晹暘傾儈僲巁攝楍傪傕偲偵堚揱巕偑扨棧偝傟偨(4)丅偙偺堚揱巕偐傜悇掕偝傟傞傾儈僲巁攝楍偼丄僸僩兝-1,4-GalT VI偲傎傏摨堦偱偁偭偨丅 丂恾1偵兝-1,4-GalT僼傽儈儕乕偵懏偡傞摐揮堏峺慺偺傾儈僲巁攝楍偺斾妑傪帵偡偑丄偄偢傟傕妶惈晹埵偱偁傞C枛抂椞堟偵崅偄憡摨惈偑尒傜傟傞丅摿偵DVD, FGGV傗GWGEDDD攝楍偺嬤朤偼丄僇僞僣儉儕偐傜偦偺堚揱巕偑僋儘乕僯儞僌偝傟偨兝-1,4-N-傾僙僠儖僌儖僐僒儈儞揮堏峺慺(兝-1,4-GlcNAcT)偵傕曐懚偝傟偰偍傝丄兝-1,4-寢崌偱摐傪揮堏偡傞峺慺偵嫟捠偡傞摐庴梕懱丄偁傞偄偼摐嫙梌懱偺堦晹(UDP)偲寢崌偡傞椞堟偱偁傞壜擻惈偑峫偊傜傟傞丅傑偨丄恾1偵帵偡4偮偺僔僗僥僀儞巆婎偺憡懳揑埵抲傕丄偙偺僼傽儈儕乕偵懏偡傞偡傋偰偺摐揮堏峺慺偱傛偔曐懚偝傟偰偄偨丅 | ||||||||||||||||
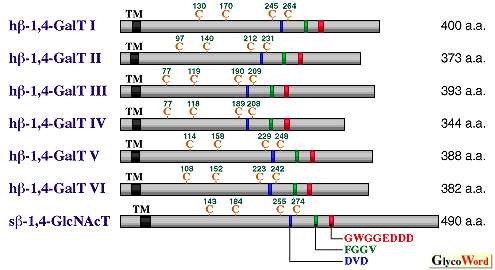 | |||||||||||||||||
| 恾侾丂兝-1,4-Gal T 僼傽儈儕乕摐揮堏峺慺偵偍偗傞傾儈僲巁攝楍偺憡摨惈 丂丂丂TM, 枌娧捠椞堟; C, 僔僗僥僀儞巆婎; h, human (僸僩); s, snail (僇僞僣儉儕) | |||||||||||||||||
|
丂兝-1,4-GalT娫偵尒傜傟傞偙偆偟偨儂儌儘僕乕偵傕偐偐傢傜偢丄婎幙摿堎惈偼奺峺慺偱堎側偭偰偍傝丄I, II, V偼摐僞儞僷僋幙摐嵔丄III偼儔僋僩-N-僱僆僥僩儔僆僔儖僙儔儈僪丄VI偼儔僋僩僔儖僙儔儈僪偺惗崌惉偵娭傢偭偰偄傞傛偆偱偁傞丅偟偐偟丄in vitro偵偍偗傞奺峺慺偺婎幙摿堎惈傛傝傕丄奺峺慺偺慻怐偱偺敪尰検傗嵶朎撪偱偺嬊嵼惈偑幚嵺偺峺慺偺婡擻偵廳梫偱偁傞偲峫偊傜傟傞丅 丂僸僩奺憻婍乮怱憻丄擼丄戀斦丄攛丄娞憻丄嬝擏丄恡憻丄鋁憻乯偵偍偗傞兝-1,4-GalT堚揱巕偺敪尰偼丄兝-1,4-GalT I, III, IV, V偼傎偲傫偳偡傋偰偺憻婍偱丄II偼怱憻丄嬝擏丄鋁憻偱丄VI偼擼偵偺傒敪尰偟偰偄傞丅傑偨丄兝-1,4-GalT I偼兛-儔僋僩傾儖僽儈儞懚嵼壓偱Glc偵Gal傪揮堏偟儔僋僩乕僗偺崌惉偵娭梌偟偰偄傞丅儅僂僗擕態偱偼兝-1,4-GalT I堚揱巕偑庡偵敪尰偟偰偍傝丄懠偺兝-1,4-GalT堚揱巕偼傎偲傫偳敪尰偟偰偄側偄丅 丂怴偟偔扨棧偝傟偨兝-1,4-GalT僼傽儈儕乕偺惗懱撪偱偺婡擻偺尋媶偼丄屄乆偺峺慺偺婎幙摿堎惈偺夝愅傕娷傔偰傑偩僗僞乕僩儔僀儞偵棫偭偨偽偐傝偱偁傝丄崱屻偺尋媶偺敪揥偑婜懸偝傟傞丅 | |||||||||||||||||
| 嵅摗丂晲巎乮搶嫗搒榁恖憤崌尋媶強丒惗懱忣曬晹栧乯 | |||||||||||||||||
| |||||||||||||||||
| 1998擭 9寧 15擔 | |||||||||||||||||
| |||||||||||||||||

