

|
|
N型糖鎖のプロセッシング異常 | |||||||||||||||||
 |
はじめに 先天性赤芽球生成異常性貧血II型、またはHEMPAS(hereditary erythroblastic multinuclearity with positive acidified serum lysis test : 酸溶血試験陽性の遺伝性多核赤芽球症)はヒトの遺伝性貧血である。正常の赤血球膜糖タンパクband 3およびband 4.5にはポリラクトサミン糖鎖が結合しているが、HEMPASではこの糖鎖を欠損している。その代わりに、糖脂質にポリラクトサミンが蓄積してくる。HEMPAS赤血球膜のN型糖鎖を分析すると一連の未熟な段階にあるN型糖鎖構造がみられ、この疾患はN型糖鎖のプロセッシング異常であると特徴づけることができる(1)。 罹患率 HEMPASは常染色体性劣性の貧血である。HEMPAS患者の両親とその兄弟の何人かは臨床症状は正常であるが欠損遺伝子をもったヘテロ接合性のキャリアーである。文献に報告されている患者数は130名であるが、300名を下らない患者がいるものと思われる。HEMPAS患者は全ての大陸に、また多くの民族に見出されている。男性女性均等に罹患する。HEMPAS患者に関する種々の症例報告によると、この疾患は多くの場合無症候性であり、それ故、今まで病気であると思う患者は非常にまれであることが示唆されている。この疾患は別の遺伝性疾患かあるいは後天性疾患に罹病した時、はじめて臨床的に顕著になるといってもよい。 症状 HEMPAS患者は一生貧血に苦しむ。通常黄疸を伴う。ほとんどの症例で貧血の程度は軽度であるが、恒常的看護と頻繁な輸液注入を要する重篤な患者もいる。HEMPAS疾患の最も直接的な診断的特徴は、骨髄において異常な多核をもった赤芽球を見ることである(図.1)。末梢血として放出された赤血球は寿命が短く、健常成人の100日に対してHEMPASのそれは7-34日である。脾臓は赤血球破壊の主要臓器であると考えられているが、HEMPASでは脾臓腫大へと移行する。脾臓の外科的切除によりしばしば貧血を軽減する。また肝では一般的に鉄沈着と肝硬変がみられる。鉄沈着と肝硬変を防止するためには、血液の抜き取りを必要とする場合もある。HEMPASの重症例では精神異常や知覚異常を示す。 | ||||||||||||||||
図.1 多核赤芽球を示すHEMPAS患者の骨髄像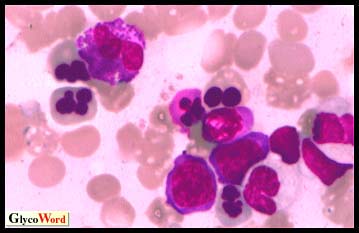
| |||||||||||||||||
|
HEMPASにおけるN型糖鎖プロセッシングの欠陥 正常赤血球では、band 3のような糖タンパクはガラクトースとN-アセチルグルコサミンの繰り返し構造から成る大きな糖鎖ポリラクトサミンによりグリコシル化されている。正常赤血球では、大部分の糖脂質は短い糖鎖を結合したグロボ系糖脂質に属している。HEMPASでは、糖タンパクはポリラクトサミンを欠き、一方ポリラクトサミンは糖脂質として蓄積する。このことはHEMPASでは遺伝的因子が糖タンパク受容体のグリコシル化を阻止し、ポリラクトサミンを脂質受容体に移行させていると思われる。HEMPAS赤血球のN型糖鎖の構造解析から、蓄積糖鎖は混成型オリゴ糖であり、N型糖鎖の生合成の崩壊はN-アセチルグルコサミントランスフェラーゼII (GnT-II) および α-マンノシダーゼ (MII) の反応ステップの周辺にあるらしいことがわかってきた(図.2)。 HEMPASの血清糖タンパクのN型糖鎖構造の解析からこれらの糖タンパクは不完全なプロセスを受けていることがわかった。このことは糖鎖生合成における欠陥は赤血球系列に限定されたものではないことを明瞭に示している。不完全なプロセスを受けているHEMPASの血清糖タンパクは肝レクチンにより認識され循環から取り除かれる。肝により取り除かれる、不完全なプロセスを受けたHEMPASの血清糖タンパクの量は非常に大量であり、このことがHEMPASにおける肝硬変の理由の一つになっていると思われる。 | |||||||||||||||||
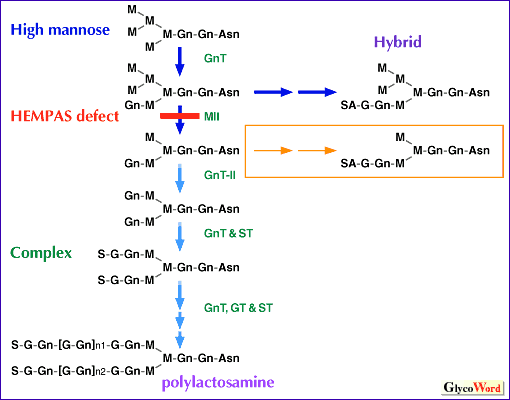
| |||||||||||||||||
|
HEMPASにおける遺伝欠陥 HEMPASは遺伝的には不均一である。MII遺伝子に欠損があると同定された最初のHEMPASの症例はイギリスからの患者である。患者のMII mRNAは正常者のレベルの10%以下しか発現していない(2)。最近、MII遺伝子をコードする領域に突然変異をもった他のHEMPASの症例が同定されている。他方、南イタリアの症例でミニSatelliteマーカーのセットを用いた糖鎖結合領域の解析からHEMPASの原因遺伝子(CDAN2)が染色体 20q11.1に同定された(3)。 CDAN2の性質は現在でもよくわかっていない。糖転移系酵素ではないあるタンパクをコードする遺伝子の欠陥がいかなる機構によりGnT-IIとMII周辺でのN型糖鎖生合成を崩壊させるのかという疑問は残ったままである。一つの可能性はゴルジ体の膜移行を制御する細胞質タンパクに欠陥が存在するということであり、この欠陥が糖鎖生合成に影響しているということである。HEMPAS患者間にみられる生化学的類似性は、不均質な遺伝子産物が生化学的に同一の経路に作用していることを示唆する。 MII遺伝子ノックアウトマウス:HEMPASの動物モデル 最近MII遺伝子を特異的に不活化した突然変異マウスが造られた(4)。MII欠損マウスは出生時は正常であり、成体へと成長する。これらのマウスには明らかな変形もなく生命をおびやかすような欠陥もみられない。しかしながらMII欠損マウスは貧血に伴う種々の兆候を示し、著しい脾臓の腫大を示す。赤血球膜タンパクのレクチンブロットから、MII欠損マウスには複合型N型糖鎖の欠失を確認できたが、これは多分N型糖鎖の生合成が高マンノース型オリゴ糖および混成型オリゴ糖へと移行したことによるものと思われる。HEMPAS赤血球に特徴的であるラクト系糖脂質の増加もMII欠損マウスに見出された。 未解決の問題点 MII欠損マウスに表れる表現形は、MIIの遺伝子の欠損とHEMPASとの間に関連のあることを支持する。しかし、MII遺伝子に本質的欠陥がおこっている症例はイギリスのHEMPASの患者に限定されていることは明らかである。南イタリアのHEMPAS患者は染色体20q11上のCDAN2にある突然変異によって罹患している。典型的なHEMPASのほかに、亜型も知られている。これらの亜型は同一遺伝子に重度の突然変異が起こることにより引き起こされた極端な症例と思われる。亜型としての表現形を示すが、未だ明らかにされていない因子がHEMPASに作用していることもあるかもしれない。MIIやCDAN2とは異なる遺伝子欠損が亜型の原因となっている可能性もある。 | |||||||||||||||||
| 福田 道子 ( The Burnham Institute ) | |||||||||||||||||
| |||||||||||||||||
| 1999年 3月 15日 | |||||||||||||||||
| |||||||||||||||||

