
Dr. Stern obtained his B.A. in 1957 from Harvard College. He worked in the laboratories of Drs. Edmond Fisher and Edwin Krebs as a medical student at the University of Washington, Seattle, where he received the M.D. degree in 1962. He was a postdoctoral fellow at the Weizmann Institute of Science, Rehovoth, Israel, in the laboratory of Dr. Uriel Littauer and then was a Research Scientist at the National Institutes of Health, Bethesda, Maryland. Since his move to the University of California, San Francisco in 1977, he has focused on the biology of hyaluronan and the hyaluronidases. He is also a board-certified Anatomic Pathologist and participates in the teaching and diagnostic services of the Department of Pathology.
Mammalian hyaluronidases were reviewed in the Science of Hyaluronan series, no. 15. More recent studies are summarized here.
Hyaluronan has an extraordinarily high rate of turnover in vertebrate tissues. A 70-kilo individual has 15 g of hyaluronan, 5 g of which turns over daily, and in the bloodstream, hyaluronan has a t1/2 of two to five minutes. Despite such dynamic degradative activity, it has previously not been possible to formulate a pathway for hyaluronan catabolism. Degradation is predominantly enzymatic, occurring in a stepwise fashion with quantum decreases in polymer size. The discrete sizes of hyaluronan fragments have widely different biological activities, indicating that a highly controlled cascade of fragmentation occurs. From these observations, a putative catabolic pathway can be formulated. The monotonous repetitive structure of hyaluronan can acquire size-dependent enzymatic substrate specificity at each stage of degradation by: 1) changes in hyaladherins with adhesion properties specific for hyaluronan polymer sizes; 2) size-dependent changes in hyaluronan molecular folding. Obviously, the two phenomena are inter-related. Most hyaladherins bind to hyaluronan by electrostatic interactions, rather than covalently, thus permitting rapid exchanges as degradation proceeds. Hyaluronidase inhibitors also exist, providing control mechanisms at various steps in the pathway. Most such hyaladherins and hyaluronidase inhibitors wait to be identified.
In the human, the three genes, HYAL1, HYAL2, and HYAL3 coding for Hyal-1, Hyal-2, and Hyal-3, are tightly clustered on chromosome 3p21.3, a region densely packed with genes. They are organized in an extraordinarily complex, and occasionally overlapping manner.1 An apparently unrelated gene, Fus2, encoding an N-acetyl-transferase, is partially imbedded in an intron of HYAL3. In the mouse, HYAL1 is often co-transcribed with HYAL3 and/or Fus2. The significance of such polycistronic transcription as well as tissue co-expression is not known, though it suggests important physiology.
From the cluster on chromosome 3p, Hyal-1 and -2 constitute the major hyaluronidases of somatic tissues. Hyal-2 is anchored to the plasma membrane by a glycosylphosphatidylinositol-(GPI-) link. Hyal-2 cleaves high molecular weight hyaluronan to a limit product of approximately 20 kDa, while Hyal-1 appears to be lysosomal, cleaving hyaluronan to small disaccharides.
Hyal-3 is a mystery. It is widely expressed, but no activity can be identified using available hyaluronidase assays. Expressions of Hyal-1, -2, and –3 occur in chondrocytes,2 and increase when fibroblasts undergo chondrocyte differentiation.3
The three genes HYAL4, PHYAL1, and SPAM1 (Sperm Adhesion Molecule1) are clustered in a similar fashion, but on chromosome 7q31.3. They code respectively for Hyal-4, a pseudogene transcribed but not translated in the human, and PH-20. This chromosomal pattern suggests two ancient gene duplications, followed by en masse block duplication of the three resulting genes.
PH-20 is the enzyme that facilitates penetration of sperm through the cumulus mass surrounding the ovum, and is also necessary for fertilization.4 It is a multifunctional enzyme protein, with a separate domain that binds to the zona pellucida.5 By more sensitive techniques involving polymerase chain reaction (PCR) analysis, PH-20 can be detected in the epididymis, seminal vesicles, prostate, 6 female genital tract ,7 breast,8 placental and fetal tissues,9 as well as in certain malignancies.8,10-12 PH-20 can also be found in renal tubular epithelia, in a form not associated with hyaluronidase activity, attributed to tissue-specific levels of glycosylation (P. Martin-DeLeon, personal communication). Thus, PH-20 is not sperm-specific, as was assumed at one time. It also indicates that these enzymes have functions in addition to their hyaluronidase activities.
The pseudogene, PHYAL1, contains an aberrant stop codon, and while not translated into active enzyme in the human, does appear to be translated in other species. This may explain why the mouse with the null mutation in the PH-20 gene is still fertile.13
A seventh hyaluronidase occurs,14 coding for an enzyme that immunologically resembles an antigen in meningiomas. The hyaluronidase activity associated with this sequence is not well characterized, but a splice variant produces a nuclear protein with β-N-acetylglucosaminidase activity.15
Tertiary structures of bacterial and vertebrate hyaluronidases are now available utilizing crystallographic analyses16 and reveal possible catalytic mechanisms. The structure of the mammalian enzymes can be modeled onto the bee venom enzyme that shares 30% sequence identity with human PH-20. The crystal structure determined at 1.6 Å resolution resembles a classical 8-fold β / α-triose phosphate isomerase (TIM) barrel, a structure common to many glycosyl hydrolases,17 except that the barrel is composed of only seven strands. A long substrate-binding groove extends perpendicularly across the C-terminal of the barrel axis. While the vertebrate hyaluronidases appear to be comprised of a combination of β-pleated sheets and α-helices in the TIM barrel motif, by contrast, the bacterial lyases or eliminase-type hyaluronidases are composed of α / α barrels. Yet they retain similar carbohydrate-binding grooves across the C-terminal.18,a
The PH-20 enzyme has a bimodal pH activity curve. The region of PH-20 encompassed by amino acid sequence 142-172 is the basis of activity at neutral pH, whereas amino acid sequence 277-297 is important for acid-active hyaluronidase. The two regions are the sites of acid/base catalysis and nucleophilic attacks respectively.19
a See article by M. Jedrzejas in this series.
It is well established that hyaluronan is taken up by cells for degradation20,21 through receptors such as CD44.22-24 The high molecular weight extracellular polymer is tethered to the cell surface by the combined efforts of receptors such as CD44 and the GPI-anchored enzyme Hyal-2. The hyluronan-CD44-Hyal-2 complex is enriched in specialized microdomains. These are invaginations of the plasma membranes composed of cholesterol and gangliosides, termed lipid rafts, significant because they also recruit a large number of key signaling molecules. One category of lipid rafts is caveolae, structures rich in the proteins caveolin and flotillin. The hyaluronan polymer is then cleaved to 20 kDa limit products,25 corresponding to about 50 disaccharide units.
The biological properties of hyaluronan in aqueous solution appear to be controlled by reversible tertiary structures, as defined by NMR spectroscopy. Evidence suggests a β-pleated sheet-like array stabilized by H- and hydrophobic bonds. Easy transitions between secondary and tertiary structures occur that are convenient mechanisms for switching between functions. The 50-disaccharide unit is around the size at which such stable tertiary structures are expected to form.26 Polymers greater than 20 kDa provide the preferred substrate for Hyal-2. The hyaladherins may provide additional substrate specificity.27 The Hyal-2-generated hyaluronan fragments are internalized, delivered to endosomes, and ultimately to lysosomes, where Hyal-1 degrades the 20 kDa fragments to small disaccharides. Two lysosomal β-exoglycosidases, β-glucuronidase and β-N-acetyl-glucosaminidase, participate in this degradation.
Evidence for the latter comes from human I-cell disease. Fibroblasts from patients with I-cell disease, lacking the mannose receptor pathway for lysosomal enzyme uptake, have an apparent hyaluronan storage disorder (Fig. 1). These fibroblasts stain intensely for hyaluronan (R. Stern and B. Steinmann, unpublished observations). The tetra- and hexasaccharide products of hyaluronan degradation are too small to be detected by the hyaluronan-binding peptide staining reaction. This suggests that the β-exoglycosidases participate actively in the degradation of 20 kDa hyaluronan fragments all along the catabolic cascade, and not only at the terminal steps.
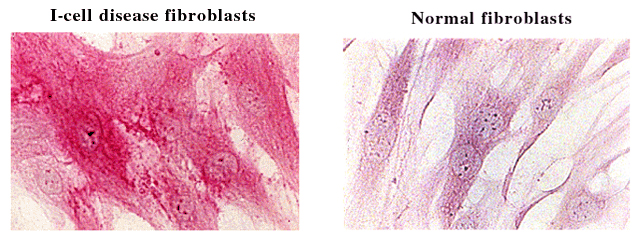
Fig. 1 Histolocalization of hyaluronan comparing cultured human fibroblasts derived from patients with I-cell disease with those from normal controls.
Cultured cells were fixed with acetone-ethanol and stained with a biotinylated HA-binding peptide derived from bovine cartilage. Cells were the kind gift of Dr. Beat Steinmann, Kinderhospital, Zürich, Switzerland.
The specific defect in I-cell disease is the enzyme N-acetylglucosamine-1-phosphotransferase, an enzyme essential for the synthesis of the mannose-6-phosphate recognition marker that targets enzymes to lysosomes. Failure of this enzyme causes misrouting of most newly synthesized lysosomal enzymes. Plasma from patients with I-cell disease have normal levels of Hyal-1, but elevated levels of the two β-exoglycosidases,28 suggesting that Hyal-1 is transported to lysosomes by a pathway different from the mannose-6-phosphate route. Without the β-exoglycosidases, larger sized hyaluronan oligosaccharides appear to accumulate in lysosomes. What may be missing is the trimming of these hyaluronan fragments to a size sufficiently small to diffuse out of lysosomes into the cytoplasmic compartment. It may be that only monosaccharides are able to diffuse out of lysosomes. Alternatively, hyaluronan fragments may leave the lysosome through specific transporters, as other metabolites do, such as amino acids and other sugars. Such putative transporters may have certain size restrictions, explaining why the larger hyaluronan fragments in I-cell disease cannot exit lysosomes. Regardless of the mechanism, it appears that oligomers that stain with the hyaluronan-binding peptide reaction accumulate in I-cell disease as a result of back-up within the lysosomes, a phenomenon that does not occur in normal fibroblasts.
An overall scheme for HA catabolism in diagrammatic form is presented below:
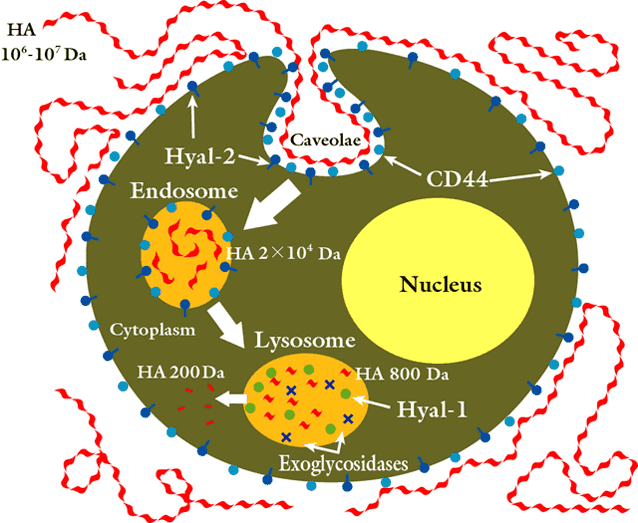
Fig. 2 Diagram of a putative metabolic scheme for hyaluronan degradation.
The hyaluronan chains can have very different biological activities, depending on chain lengths. High and low molecular weight chains appear to have opposite and opposing effects on cell behavior. The extracellular high molecular weight (~107 Da) hyaluronan chains are space-filling molecules that hydrate tissues, and inhibit angiogenesis.29 They are also anti-inflammatory, and immune-suppressive.30,31 The low molecular weight hyaluronan fragments interact with a different set of receptors that trigger signaling cascades and profound changes in cell behavior. Among the functions of the large polymers, then, is to protect the cell from access to such fragments, to impede cell-cell interactions, and to inhibit stress responses and signaling cascades.
High molecular weight hyaluronan appears in the earliest stages of wound healing, when spaces must be created to facilitate polymorphonuclear leukocytes infiltration of the wound area. These cells clean the wound site, remove debris, dead tissue, and engulf bacteria. The same high molecular weight hyaluronan polymers have an increased ability to bind fibrinogen,32 one of the first reactions to occur in clot formation, critical in early wound healing.33
The 20 kDa limit fragments of Hyal-2 digestion are cleaved to intermediate-sized highly angiogenic, immune-stimulating fragments. They are also potent stimulators of inflammatory cytokines.34 Hyaluronan is found in high molecular weight form in the normal lung, but following acute lung injury, hyaluronan occurs as chains smaller than 500 kDa, at concentrations that can initiate and maintain inflammatory pathways (M.M. Mascarenhas, personal communication).35 Smaller hyaluronan oligomers, in the 6-20 kDa size range, are potent activators of dendritic cells, the antigen presenting cells of the immune system.36,37
Very small hyaluronan oligosaccharides also have unique biological activities. Oligomers of six disaccharides promote differentiation of the endothelial cells induced in response to the angiogenic affects of larger hyaluronan fragments, suggesting a carefully controlled cascade in the course of hyaluronan degradation. Controls of migration, maturation, and signaling in skin keratinocytes are also closely associated with small hyaluronan oligomers, with widely different signaling pathways elicited by changes in polymer size (K. Törrönen et al., personal communication). The exquisite selectivity of the system is reflected in the observation that hyaluronan bound to CD44 on keratinocytes can be displaced by hyaluronan decasaccharides, but not by hexasaccharides.38
Tetrasaccharides are among the predominant products of Hyal-1 digestion. These, when added to cultured cells, induce expression of heat shock proteins39. The tetrasaccharides are also anti-apoptotic, suppressing cell death in cultures undergoing hyperthermia or when cells are serum-starved.39
Hyal-1 is a candidate tumor suppressor gene product, deleted in many tobacco-related tumors, not only of the lungs,40 but also of the head, neck, and upper airways.41 The enzyme can be eliminated at the level of DNA, by homozygous deletion or by loss of heterozygosity, but also at the level of RNA. Several studies have documented that two transcripts occur, the larger transcript containing a retained intron that prevents translation.42,43
Hyal-2 can function as either an oncogene or a tumor suppressor gene product. Over expression of Hyal-2 accelerates tumor formation of murine astrocytoma cells.44 Hyal-2 is also the cell surface receptor for some retroviruses, the envelope proteins of which mediate oncogenic transformation.45-47 Evidence for tumor suppressor gene function of Hyal-2 is also available. Hyal-2 can accelerate apoptosis.48 Furthermore, an adenovirus-Hyal-2 vector suppresses growth of tumor xenografts in mice.49 Finally, Hyal-2-over-expressing clones of src-transformed fibroblasts have reduced rates of proliferation (B. Flamion, personal communication). The conundrum that both hyaluronan and hyaluronidases are involved in tumor promotion as well as in tumor suppression will be addressed in another communication.50
Small hyaluronan oligosaccharides induce cleavage of CD44 at the surface of cancer cells that promote motility and invasion. The hyaluronidase activity generating these 6 to 14 disaccharide unit fragments appears to be a cell-surface activity that does not occur prominently in normal cells.51 Intricate enzymatic modifications occur on the surface of cancer cells including enhancement of proteases such as matrix-metalloproteases and cathepsin B. Such modifications apparently include hyaluronidase activities.
Hyaluronan oligomers have the ability to suppress tumor growth. This appears to occur by multiple mechanisms. Hyaluronan oligomers can inhibit tumor formation in vivo. Injected subcutaneously, they inhibit tumor growth, perhaps by competing with high molecular weight matrix HA for CD44. This prevents the adhesion necessary for tumor growth and invasion.52 This inhibition parallels the ability of soluble CD44 to also prevent the growth of tumor cells.53 Hyaluronan oligosaccharides inhibit anchorage-independent growth of tumor cells by suppressing the phosphoinositide 3-kinase/Akt cell survival pathway,54 as well as inhibiting multidrug resistance.55 The HA fragments thus have intrinsic tumor suppressive effects on tumor growth as well as sensitizing cells to chemotherapeutic agents.
The caveat is that cancer cells do things differently, and that the catabolic scheme described here may require modifications when applied to malignant cells.
This formulation however has a number of provisos: 1) Products of hyaluronan synthase 3 (HAS3) are small hyaluronan fragments, compared to the high molecular weight products of HAS1 and HAS2. Levels of HAS3 are increased under stress situations such as in acute lung disease.35 Therefore inflammatory hyaluronan oligomers are generated not only from products of hyaluronan degradation, but under stress situations, can also be direct products of hyaluronan synthesis; 2) Fragmentation of hyaluronan can also occur in the presence of free radicals under oxidative conditions from reactive oxygen intermediates that degrade hyaluronan by random cleavage of endoglycosidic bonds.56,57 Free radicals and hyaluronidases may have coordinated hyaluronan chain scission activities under certain pathologic conditions, such as in the respiratory burst of polymorphonuclear leucocytes that accumulate in the first stages of inflammation and early in wound healing.58 The relative proportions of free radical scission and enzymatic degradation have not been quantified, under either normal or pathologic conditions; 3) Many of the observations used in the formulation of this catabolic scheme are derived from experiments in which sized hyaluronan oligomers are added to cultured cells. It is not at all clear whether this represents a physiological situation. Addition of hyaluronan fragments exogenously to cultured cells may not be equivalent to the generation intracellularly of similar hyaluronan fragments. How intracellular hyaluronan oligomers exit cells, if they exit at all, has also not been established; and 4) Contaminants in hyaluronan preparations may be the source of the pro-inflammatory activity. Low levels of DNA in commercial preparations are suspected to be the source.59 Highly purified hyluronan is non-inflammatory across a broad range of molecular weights (Hu et al., personal communication). If this phenomenon is verified, that low molecular weight fragments of hyaluronan are not the basis of the angiogenesis and inflammation, a reevaluation of the pathobiology of the catabolic scheme will be necessary.
There are many unanswered questions in a putative scheme for hyaluronan turnover and degradation, particularly in the control mechanisms involved. A tentative catabolic scheme is presented here, in which step-wise degradation of hyaluronan is described utilizing a series of sequential enzyme reactions. Elevated levels of particular fragments might occur in association with specific pathologic states. Stimulation of Hyal-2 activity, or inhibition of Hyal-1 would cause inflammatory, immune-stimulating and angiogenic fragments of hyaluronan to accumulate.
Hyaluronidase inhibitors would provide rapid response elements, and may reflect why turnover rates are so very rapid. Release from constant inhibition would provide the mechanism for prompt increases in hyaluronan levels. Such inhibitors have been detected in many tissues. This is supported by indirect evidence, by the increase in apparent levels of total hyaluronidase activity following the initial steps in enzyme purifications from liver, muscle, spleen, heart (unpublished observations), and serum,60 as inhibitors become separated away from enzymes. Such inhibitors may be more diverse than the hyaluronidases themselves.61,62
Characterizing all aspects of the catabolic pathway for hyaluronan becomes an enormously complex task. The particular hyaladherins, and specific inhibitors associated with the hyaluronidases and their hyaluronan substrates at each step will have to be determined. The elucidation of the catabolism of the monotonously repeating hyaluronan sugar chain may be far more arduous than that of the more complex proteoglycan-associated glycosaminoglycans.



