
Davide Vigetti
Davide Vigetti received his degree in biology cum laude in 1996 at the University of Milan (Italy). Then he moved at the Faculty of Sciences of the University of Insubria (Varese, Italy) and obtained a PhD in Evolutionary and Developmental Biology in 2001 studying the molecular mechanisms of the progressive loss of the purine degradation pathway in vertebrates taking the amphibian Xenopus laevis as a model. He became an Assistant Professor of Biochemistry in 2004 at the School of Medicine of the University of Insubria, where he discovered the amazing world of glycobiology and started to study the intracellular mechanisms that control hyaluronan synthesis identifying the critical role that UDP-sugar precursors have not only in Xenopus but also in mammalian cells.

Michela Ori
Michela Ori is a Young Group Leader in the Unit of Cellular and Developmental Biology, Department of Biology, University of Pisa, Italy. She received her PhD in “Molecular Biology and Biophysics of the Cell and Development” at the University of Pisa, in January 2001 working on the role of the homeotic gene Hoxa2 during cranial neural crest cell development using the Xenopus as a model system. During this project she spent one year in Prof. F. M. Rijli’s laboratory at the IGBMC, Strasburg, France. After returning to Pisa she maintained an efficient collaboration with the group of Prof. F. M. Rijli studying the genetic control of neural crest cell development in Xenopus and started new projects in the field of developmental neurobiology. The interest in developmental biology raised the question of how cells interact with each other and with the extracellular environment. In particular, thanks to collaboration with Prof. R. Perris of the University of Parma, Italy, Michela Ori started a new project on the role of hyaluronan during Xenopus development. From 2005 she established her own lab now focusing on the role of hyaluronan, hyaluronan receptors and versican in neural crest cell development and in muscle regeneration. The frog is still our beloved model system!

Alberto Passi
Alberto Passi MD PhD is full professor in biochemistry at the Medical School, University of Insubria (Varese – Italy), dean of the Faculty of Exercise and Sport Sciences of University of Insubria, director of biochemistry laboratories at Department of Biomedical Experimental and Clinical Sciences (DSBSC) of University of Insubria. Since obtaining his PhD, he has focused his scientific interest in extracellular matrix metabolism. Cell cultures, lung and vascular tissues were initially approached studying proteoglycan metabolism including the effect of free radicals and proteases on extracellular matrix macromolecules. After a sabbatical period in Hascall’s lab in Cleveland (OH – USA), the Passi work moved towards hyaluronan. In particular, studies were done on hyaluronan metabolism and its control. The role of UDP sugars precursors in glycosaminoglycan synthesis and the covalent modification of enzymes involved in glycosaminoglycan metabolism are currently the main interest in Passi’s lab.
Hyaluronan (HA) is an abundant glycosaminoglycan (GAG) of the embryonic extracellular matrix (ECM) with critical biological functions. Its importance is recognized for the correct assembly of the interstitial matrices and for its putative ability to influence cell behaviour through binding to signal transducing receptors (i.e., CD44, RHAMM, TLR4/2, LYVE-1 and HARE). In this review we describe the importance of HA and of enzymes involved in its metabolism in the amphibian Xenopus laevis, a well known model organism to study vertebrate development. Moreover, as ontogenesis recapitulates phylogenesis, we discuss some evolutional issues about HA and other GAGs.
The ECM has multiple roles in a living organism. It has structural, mechanical, and trophic functions permitting the exchange of nutrients and wastes. Moreover, ECM is the “battlefield” of immune cells that migrate from the vessels to tissues to fight against pathogens or to remove the remains of physiological cell turnover. In addition, ECM can bind signal molecules like growth factors, mitogens, chemokines and cytokines, which makes the tissue microenvironment suitable for cell proliferation, migration, and differentiation. During embryonic development, morphogen gradients are maintained through specific ECM components that permit the correct series of spatial-temporal signaling events. The ECM composition can vary to meet functions of the considered tissue or cell. Moreover, it is widely accepted that ECM remodeling is a critical step in the response to both physiological and pathological stimuli.
The presence of polysaccharide components of ECM is conserved throughout evolution from unicellular to pluricellular organisms, and a major reason for the presence of sugars is their huge variability that permits specific cell recognition and interaction. In vertebrates, the main ECM polysaccharides are HA, chondroitin sulfates, dermatan sulfates, heparan sulfates and keratan sulfates, which, with the exclusion of HA that is not normally covalently bound to a protein, constitute the GAG chains of proteoglycans (PGs).
HA is a linear unsulfated GAG that is composed of repeating units of D-glucuronic acid and N-acetylglucosamine linked together through alternating beta-1,4 and beta-1,3 glycosidic bonds. In contrast to the other GAGs that are synthesized in the Golgi, HA is polymerized directly on the plasma membrane by a family of three HA synthases (HAS 1, 2, and 3) that use cytosolic UDP-GlcUA and UDP-GlcNAc as substrates 1. These latter compounds are synthesized by UDP-glucose pyrophosphorylase (UGPP) and UDP-glucose dehydrogenase (UGDH), which work in sequence in the cytoplasm. The length of the polymeric chain can vary greatly because HASs have different biochemical properties 2, and also because several isoforms of HA degrading enzymes, hyaluronidases (Hyal), exist 3.
The relatively simple chemical structure of HA lacks the structural complexity of the other GAGs due to their sulfation and the presence of core proteins in PGs. Therefore, the only way to introduce information of biological relevance for HA is the variation of its size as well as the structure of the proteins that interact with HA. Hence, the molecular mass strongly influences the effect that HA has on cellular properties 4. In this context, it is clear that the HA content in the amphibian Xenopus laevis, a model organism to study development in vertebrates, can modulate cell behavior, and this topic is the main aim of this review. Moreover, as ontogeny recapitulates phylogeny, we discuss possible differences in the evolution of HA and the other GAGs.
GAG assembly strongly depends on the availability of UDP sugars in the cells 5. This aspect of cell biosynthesis implies that cell metabolism supports the anabolic pathways, and therefore the cells producing GAGs must have a positive energy balance. In fact, UDP sugar precursors represent molecules with a high cost in terms of energy, as UDP sugars compete with glycolysis for their synthesis. For this reason, it is evident that GAG synthesis is easier in healthy tissues with a good oxygen supply, as these tissues down regulate glycolysis while the presence of oxygen allows the oxidative reaction necessary for UDP-GlcUA synthesis (Fig. 1). The synthesis of UDP-GlcUA is a critical step for all GAGs, except keratan sulfate (KS), which does not contain uronic acid. Further, KS is present primarily in tissues without a vascular system and consequently with a poor oxygen supply, such as cornea and the inner portion of articular cartilage 6, 7. It was suggested that in these areas, KS may represent a biological substituent for CS, which requires the uronic acid for its synthesis and therefore a high oxygen supply 8. It is not surprising that KS is almost absent in fetal tissue, whereas it increases in tissues characterized by low oxygen supply and during tissue aging.
The synthesis of UDP-GlcUA requires the activity of the UDP-glucose dehydrogenase (UGDH), which produces UDP-GlcUA from precursor UDP-Glc. This reaction is possible in the presence of NAD, which is converted in NADH during the two fold oxidation of the C-6 of UDP-Glc. This reaction is quite unusual. A double oxidation on the same C-6, which is converted from alcoholic to carboxylic group producing two NADH/mol of UDP-Glc, is very uncommon in nature. Moreover, this aspect of GAG synthesis introduces the importance of UDP-GlcUA in terms of energy balance. When all the steps of the anabolic pathway from glucose to UDP-GlcUA are considered (Fig. 1), it is evident that the costs of UDP-GlcUA synthesis are completely repaid by the reoxidation in mitochondria of NADH to produce ATP molecules. In fact, considering the stoichometry of GAGs, which are characterized by the presence of UA and hexosamine in a ratio 1:1, the energy cost of the synthesis of the unsulfated backbone of GAGs is completely sustained by C-6 oxidation of UDP-Glc. This is particularly remarkable for HA, where the absence of sulfation reduces the energy costs to the GAG backbone, and the energy balance to synthesize this GAG is completely compensated by the reoxidation of the NADH produced in UDP-GlcUA synthesis.
We investigated the availability of UDP sugars precursors on GAG synthesis and demonstrated that alteration of the UDP-GlcUA concentration in the cells influences HA synthesis and also modifies the gene expression pattern of enzymes involved in its synthesis. This indicates the presence of cross-talk between the cytoplasm and nuclei 5.
Interestingly, HA appeared relatively late in evolution with its synthesis appearing to start from the early chordates, although some microbial pathogens can synthesize HA to elude the host immune system. However, they likely acquired their HAS enzymes from their hosts since their HAS are homologous to the mammalian HASes although the origin of their HA synthetic pathway is still debated (see the Hyaluronan Today review of Weigel for a more complete discussion about this topic).
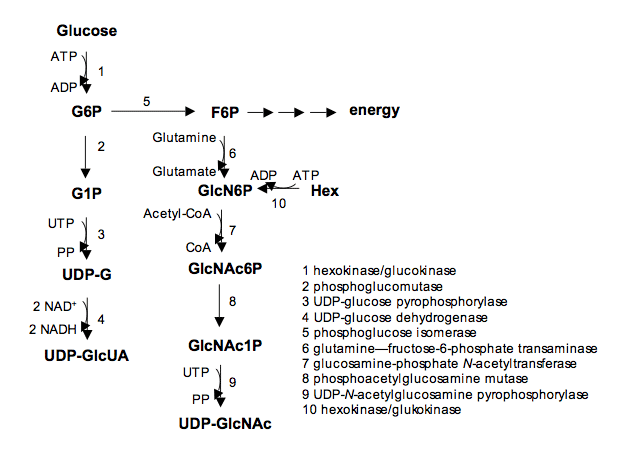
Fig. 1
Intracellular biochemical pathway of the biosynthesis of HA precursors. G6P, glucose 6 phosphate, G1P, glucose 1 phosphate, UDP-G, UDP glucose, UDP-GlcUA, UDP-glucuronic acid, F6P, fructose-6-phosphate, GlcN6P, glucosamine-6-phosphate, GlcNAc6P, N-acetylglucosamine-6-phosphate, GlcNAc1P, N-acetylglucosamine-1-phosphate, UDP-GlcNAc, UDP-N-acetylglucosamine. Hex, hexoses. Each biochemical reaction is catalyzed by a specific enzyme reported as numbered.
GAGs, like other ECM components, turn over in tissues. The coordinated action of Hyals outside the cells as well as inside the lysosome together with beta-glucuronidase and hexosaminidase lead to the formation of free hexosamines and GlcUA. While hexosamines are rapidly recycled in the hexosamine biosynthetic pathway producing UDP-hexosamines and, in turn, new polysaccharides (Fig. 2) 9. The fate of GlcUA has more metabolic significance, and only in plants and probably in some microorganisms does a clear recycling of GlcUA occur. Further, the direct phosphorylation of GlcUA to GlcUA -1- phosphate (GlcUA-1P) and the conversion of GlcUA-1P to UDP-GlcUA only occurs in such organisms 10. In vertebrates, GlcUA enters into a series of redox and decarboxylation reactions that converts it into D-xylulose-5-phosphate, which is an intermediate of the pentose phosphate pathway (Fig. 2) 11. In terms of energy balance, the pentose final product reflects the initial oxidation of UDP-Glc operated by UGDH. In fact, the decarboxylation from 3-keto-gluconate to D-xylose eliminates the completely oxidated C-6 product in the anabolic process. This energetic aspect of HA metabolism underlines a sophisticated mechanism by which cells are able to obtain energy during the biosynthetic process of an extracellular polymer, and how the final balance is compensated by the recycling of the uronic moieties in the pentose pathway. Considering the massive turnover of HA in vertebrates, it is not surprising that the pathway for recycling of uronic acid is involved in an important human metabolic disease called pentosuria, caused by l-xylulose reductase deficiency. Affected patients are characterized by abnormal amounts of l-xylulose in the urine but without any severe complications.
Interestingly, GlcUA is the starting substrate of vitamin C biosynthesis, a pathway that occurs in most vertebrates, although not in guinea pigs and primates, including humans. However, it seems that the GlcUA for vitamin C synthesis derives from intracellular breakdown of UDP-GlcUA, and no literature links GAGs turnover and vitamin C 12. On the contrary, myo-inositol, which is considered a vitamin for certain vertebrates, like mice and rats but not for humans, is degraded to GlcUA by the reaction catalyzed by myo-inositol oxygenase.
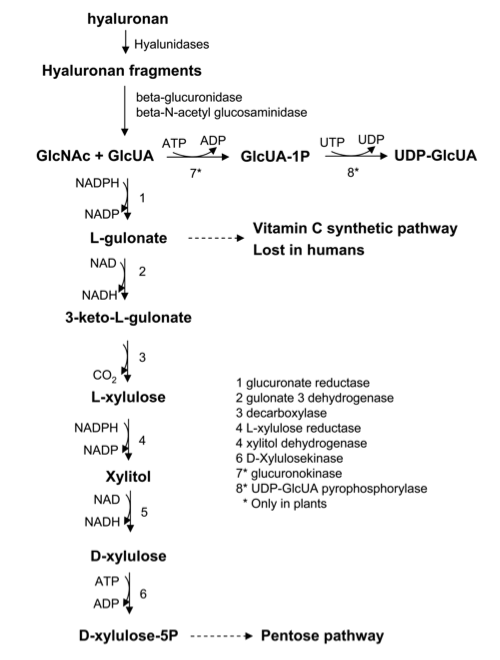
Fig. 2
Biochemical pathway of HA degradation. GlcUA-1P, glucuronate-1-phosphate. Each biochemical reaction is catalyzed by a specific enzyme reported as numbered. Note that reactions 7 and 8 are present only in plants and some microorganisms.
While researchers might argue the advantages of their particular model system, it is important to realize that complementary experimental approaches in different systems have greatly increased our progress in understanding the overall processes occurring during embryogenesis.
For many decades Amphibia were a favorite model system of embryologists due to the large size of the eggs, the large number of embryos, which develop outside of the mother, and the fact that the embryos exhibit a remarkable ability to heal after microsurgery. One disadvantage of traditional amphibian species is that they are seasonal breeders. This meant that investigators could not do their experiments throughout the year. Xenopus laevis, the South African clawed frog, is a notable exception. Its ability to spawn when injected with gonadotropic hormone led to its common usage for human pregnancy tests in the 1950s where an injection of pregnancy urine, which contains chorionic gonadotropin, would induce spawning. This led investigators to consider its use in experimental embryology.
Xenopus embryos develop outside of the mother, which makes themextremely well suited for manipulations of gene activity via microinjection. Typically microinjection studies are used to examine the function of a gene of interest by overexpression of the wild type, or a mutant sequence of the gene, or by loss of function approaches. Furthermore, the extremely reproducible cleavage patterns of the frog embryo and the availability of detailed fate mapping studies facilitate targeting of injected material to restricted lineages of the developing embryo.
A complementary approach of gene loss of function can be done by microinjection of antisense oligonucleotides into the Xenopus embryo to provide gene-specific inhibition of expression. While numerous types of modified antisense oligonucleotides exist 13, morpholino oligomers (MOs) are non-toxic, stable and highly efficient. Therefore, they have become widely used by Xenopus researchers. MOs knockdown the expression levels of a target gene by interfering with either translation of the target mRNA or by splicing of the target pre-mRNA. The resistance of MOs to endogenous nucleases results in efficient and sustained reduction of the targeted message within the injected cell and in all of its progeny. A gradual loss of activity occurs at later stages of development, which may be due to dilution 14. The Xenopus life cycle and the microinjection technique are described in Fig. 3.
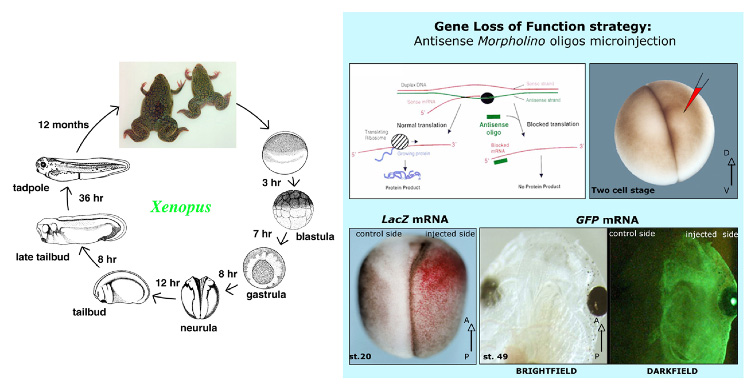
Fig. 3
On the left, the life cycle of Xenopus laevis. On the right, the gene loss of function strategy that specifically blocks the translation of a gene of interest by microinjecting specific morpholino oligos in one side of the embryo. The injected side of the embryo is visualized by the co-injection of a reporter gene such as the LacZ or GFP. An example of chromogenic reaction for beta galactosidase is shown in red in a neural stage embryo, while the green fluorescence reveals the injected side in a living larva embryo.
UDP-glucose dehydrogenase (UGDH) supplies the cell with UDP-GlcUA, a substrate for the synthesis of all GAGs with the exception of keratan sulfate. In addition, UDP-GlcUA is used in the liver where glucuronidation targets molecules for excretion. UDP-GlcUA also serves as a precursor for UDP-xylose, which provides a major component of cell wall polysaccharides in plants and provides the initial sugar in GAG synthesis on the core proteins of most PGs. Because of the central role of UDP-GlcUA, it is not surprising that alteration in UGDH expression causes evident phenotypes in developing embryos. In Drosophila melanogaster UGDH is encoded by the sugarless gene and is required for heparan sulfate synthesis on PGs that control wing formation 15. In Caenorhabditis elegans, UGDH influences GAG synthesis, which is essential for vulval morphogenesis and embryonic development 16, 17. In Zebrafish, the enzyme is critical for normal cardiac development 18. In mouse, UGDH mutants arrest growth during gastrulation with defects in migration of cells in mesoderm and endoderm 19. In Xenopus, UGDH is expressed throughout the entire development with a peak in the tailbud stage 5. The high UGDH expression is not surprising as it is known that HA and the GAGs of PGs are essential for proper development. Interestingly, the timing of the peak of UGDH expression correlates with the peak of HA synthesis during embryogenesis (Fig. 4B, 32 hours post fertilization (HPF)).
UDGH has a central role in HA metabolism by providing a substrate of HASs. Moreover, the concentrations of UDP-sugars, in particular UDG-GlcUA and UDP-GlcNAc, have a critical role in modulating HA synthesis 5. In contrast, the synthesis of other GAGs was not affected by changes in the UDP-GlcUA pool suggesting that the mechanism of the UDP-sugar antiport transporters can maintain a constant concentration of the UDP-sugars inside theGolgi compartment where other GAGs are synthesized.
The role of UGDH in Xenopus development was demonstrated by loss of function experiments with injection of morpholino oligos. Embryos treated with an MO against UGDH showed severe malformations at the end of gastrulation that led to a failure in the blastopore closure and to a premature death before the tail bud stage. During gastrulation, both in Xenopus laevis and mouse embryos, mesoderm cells enter and move within an HA-rich extracellular matrix environment 20, 21. The inhibition of synthesis of this HA-rich ECM by UGDH silencing impaired critical cell movements that form a normal gastrula constituted of three embryonic germ layers: ectoderm, mesoderm and endoderm.
HA is absent in oviposed eggs and in their coats (i.e., vitelline envelope and jelly layers). In keeping with the absence of the transcripts of XHAS1, 2, and 3 in eggs. HA levels increase in stages corresponding to late gastrulation, neurulation and organogenesis during early development (Fig. 4). The increase of XHAS1 transcript followed by increases of XHAS2 and 3 correlates well with the HA peak, which reaches its maximum at 32 h post fertilization (Fig. 4B) and with increasing activities of HA synthesis detectable in X. laevis embryos extracts 22. The rapid decrease that follows correlates with the appearance of the XEH1 hyaluronidase and an increase of HYAL2.
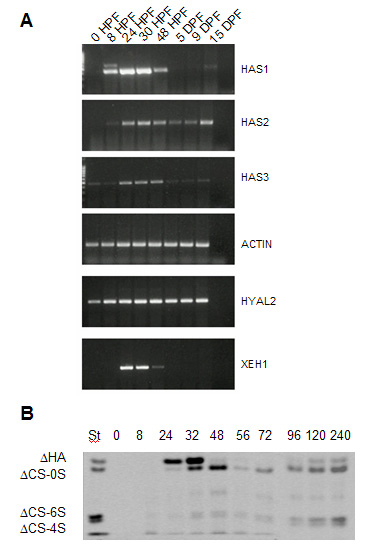
Fig. 4
A. Expression of HA metabolizing enzymes during Xenopus laevis development by RT-PCR.
B. Quantification of HA during early phases of Xenopus laevis development by PAGEFS. These images are included in Vigetti et al. (2003) and reproduced with permission of Elsevier.
The spatiotemporal distribution of HA showed that it is present in the blastocoel, and the archenteron, as well as later on in the hepatic cavity, the brain ventricles and the developing heart. At the blastula stage, HA is detected in the extracellular matrix of the ecto- and mesodermal primordial, whereas during gastrulation, HA is in the involuting marginal zone and in the deep layer of the equatorial mesodermal primordium. After gastrulation, HA appears to accumulate within the extracellular matrix demarcating the primary germ layers and then in many mesodermal derivatives, e.g., in mesenchyme, heart, prechordal cartilage and the lung primordial 21.
Vertebrate HA biosynthesis is governed by three catalytic enzymes that reside at the plasma membrane and are designated Has1, Has2, and Has3. The HAS1 gene was isolated in 1983 23 together with other genes that are Differentially expressed in developing Xenopus at Gastrulation and given the name DG42. DG42 represents the first discovered eukaryotic HA synthase although its biochemical activity was debated between HA or chitin synthase for some time 24. The three synthases are expressed in different temporal patterns during mouse development 25, and in particular, Has2 has been identified as a major source of HA during initial organogenesis. Mice with a homozygous deletion of the Has2 gene manifest severe cardiac and vascular abnormalities leading to death at midgestation (E9.5-10), underpinning a pivotal role of Has2 during mammalian embryogenesis 26. The technical limitations in carrying out detailed analyses of the Has2 gene function by transgenic approaches in rodents have led to analogous investigations in more accessible embryos, such as that of the fish and frog. Down regulation of the Has2 gene in zebrafish perturbs the mesodermal cell movements responsible for the gastrulation process and the proper heart formation 27, 28.
Overexpression of Has2 in the mesoderm of the chick limb bud in vivo results in the formation of shortened and severely malformed limbs that lack one or more skeletal elements. Skeletal elements that do form in limbs overexpressing Has2 are reduced in length, exhibit abnormal morphology, and are positioned inappropriately. It has also been demonstrated that sustained HA production in micromass cultures of limb mesenchymal cells inhibits formation of precartilage condensations and subsequent chondrogenesis, indicating that downregulation of HA is indeed necessary for formation of the precartilage condensations that trigger cartilage differentiation. Taken together these results suggest involvement of HA in various aspects of limb morphogenesis 29.
We and others, have recently cloned the three Has genes in Xenopus laevis and established their spatio-temporal distribution during early phases of development 20, 30, 31. XHas1, XHas2 and XHas3 mRNAs exhibit distinct and non-overlapping spatial expression patterns (Fig. 5) that differ significantly from those observed in zebrafish, and which suggest an evolutionary divergence in the control of Has genes expression between tetrapods and teleosts. In Xenopus, expression of XHas1 and XHas2 is first detected in the animal pole of midblastula embryos. Starting from the early gastrula stage, XHas1 mRNA is localized within the entire ectoderm while XHas2 is expressed in the involuting mesoderm. At early neurula, XHas1 is absent from the neural tube, but a signal becomes evident in pre-migratory cranial neural crest cells. At the neurula stage, XHas2 is expressed within presomitic mesoderm, and at the tailbud stage, the expression of XHas2 is evident in the segmented paraxial mesoderm. In the same stages XHas2 mRNA is detectable in the trunk neural crest cells and in the heart primordium. On the other hand, XHas1 expression becomes down regulated in the ectoderm following a dorso-to-ventral sequence across the embryo. In the head region, XHas2 mRNA is present in the head mesenchyme and in an anterior region containing the pituitary anlage. XHas2 also appears in the eye lens and in the branchial arches.
XHas2 expression follows the myogenic wave of segmentation of the paraxial mesoderm along the antero-posterior axis. The XHas2 expression, in fact, diminishes in the differentiated anterior myotomes, remaining present only in the posterior somites and in the paraxial mesoderm of the tailbud. Whereas XHas1 and 2 are widely expressed in the embryo, the transcription of XHas3 is restricted to certain areas of the embryo, including the inner ear and the cement gland. In conclusion this comparative gene expression analysis suggests that the different roles hyaluronan has during development could be modulated by the spatio-temporal regulated transcription of the three different synthases.
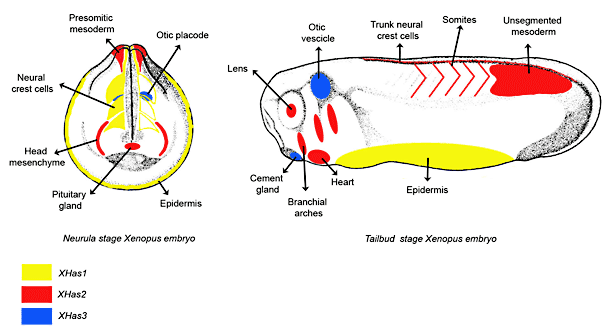
Fig. 5
Schematic representation for the gene expression patterns of the three HA synthases during Xenopus embryo development.
The early lethality of Has2 knock out mouse embryos leaves the precise role of HA during embryogenesis partially unresolved. To approach this problem we pursued a loss of function study that allows us to delineate the XHas2 role in early developmental processes 32. Tailbud Xenopus embryos that received a unilateral injection of XHas2 morpholino at the two-cell stage failed to correctly organize their flank musculature. The embryos had smaller somites, and the segmental organization of the myotomes appeared disrupted. These observations raised the possibility that XHas2 loss of function could affect the structural integrity of the inter- and intra-somitic extracellular matrix, which in turn could interfere with the establishment of correct cell-cell contacts between the forming myoblasts.
As XHas2 is expressed in presomitic cells, we investigated whether the loss of Has2 activity actually interferes with the early phases of the myogenic differentiation program. At the neurula stage, we observed a significant increase in the number of apoptotic cells in the presomitic mesoderm while myoblast proliferation and the expression of the precocious muscle markers appeared unaffected. These data strongly suggested that XHas2 activity is not essential for the early myogenic commitment or myoblast proliferation, but could be required for the survival and differentiation of the early muscle precursors. Indeed, the role of XHas2 in preventing these cells from undergoing apoptosis seems to be restricted to a specific developmental period, which coincides with the early steps of myogenic differentiation in the presomitic mesoderm. Proper assembly of the mesodermal ECM in the presence of HA may be important for the maintenance of community effects, for proper diffusion/gradient formation of morphogens, or for undisturbed action of other instructive signals involved in the differentiation and survival of developing myoblasts.
The effect of HA on survival and differentiation of presomitic mesodermal cells could be exerted through a structural supportive effect of the ECM. A more intriguing possibility is that HA produced by myoblast cells could activate intracellular signals via binding to specific receptors such as CD44 or RHAMM. To further investigate this aspect, we performed functional knock down of XCD44. In Xenopus this receptor is expressed in the developing musculature, but its deletion did not produce significant alteration in the formation of trunk muscle. Conversely, both XHas2 and XCD44 are involved in hypaxial myoblast migration, which in Xenopus is required for the proper formation of the ventral body wall musculature. The expression of XHas2 and XCD44 in this precursor cell population suggests that the impaired migration may, at least in part, be determined by the failure of hypaxial cells to contribute with their proper HA during the migratory process and that XCD44 function may be required for cell locomotion. It has been shown in zebrafish that Has2 is required to activate Rac1 signalling cascades that govern dorsal cell migration during gastrulation and adaxial cell migration in the forming somites, allowing for the hypothesis of an HA instructive role in inducing cells to migrate 27. We further suggest that, at least for hypaxial muscle cells, this HA role may be mediated by XCD44 activation 32. Another interesting phenotype, observed in XHas2-deprived embryos, is an altered trunk NCC migration, corroborating a proposed pivotal role of HA during NCC development 33. It is noteworthy that Xenopus migrating trunk NCCs do not express XCD44, and that depletion of the receptor in somitic cells does not influence the NCC movements. Although the presence of HA in the extracellular environment seems to be crucial for allowing proper NCC migration, it is not clear if the impaired movement of trunk NCCs observed in XHas2-depleted embryos was due to a failure of these cells to synthesize and secrete HA, or to a structural deficit caused by its loss from the ECM of the NCC migratory pathways. An alternative possibility could be that the altered NCC migration could result from a morphogenetic loss of the structural integrity of the somites through which the cells migrate. Further studies are needed to fully clarify this issue, but these preliminary results may suggest that both a correct structure of somite and a HA enrichment of the ECM, are necessary to create a specific environmental condition necessary to allow NCC migration 32.
We are now exploring the possibility of alternative molecular mechanisms of action of HA during development studying possible functional interactions of HA with alternative receptors, such as RHAMM, and hyaluronan binding partners such as the proteoglycan versican 34.
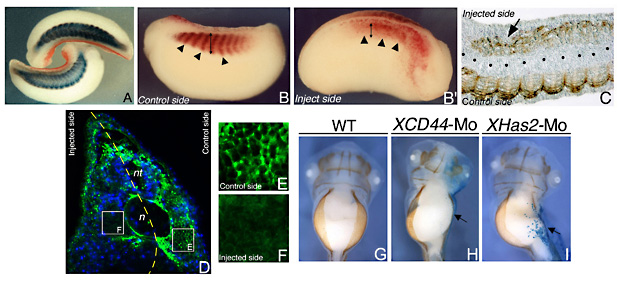
Fig. 6
Myotome alterations in XHas2-Mo injected tailbud embryos. (A) Embryos bend towards the injected side as visualized by the red-gal staining. Somites are visualized by the mRNA expression of cardiac actin in blue. (B, B, C) 12/101 muscle specific antibody immunoreactivity (brown staining). (B) Lateral view of the control side of a stage 26 embryo (arrowheads indicate the somites). (B) Injected side of the embryo in (B) showing a complete disruption of somite segmentation as indicated by the arrowheads, and a strong reduction in the 12/101 positive cells (compare arrows in B and B). (C) XHas2-Mo injected tailbud embryo longitudinally sectioned. Note the altered pattern of the 12/101 positive myocytes within the somites in the injected side (arrow). (D) Coronal sections of an XHas2-Mo injected embryo at the trunk level were stained with neurocan-GFP fusion protein in green, and nuclei in blue. (E) High magnification of the control side showing abundant HA in the ECM in the myocyte extracellular spaces. (F) High magnification of the XHas2-Mo injected side. No HA detection is visible in the ECM surrounding the myocytes.
(G, H, I) Ventral views of stage 43 embryos immunostained with the 12/101 antibody that shows the final position of the ventral body wall musculature in wild type (G), XCD44-Mo injected embryo (H) and XHas2-Mo injected embryo (I). The side of injection is visualized by X-gal staining in blue and indicated by the arrow. n, notochord, nt, neural tube.
These images are included in 32 and reproduced with permission of the Company of Biologists.
The cell within its extracellular matrix, represents a morphogenetic unit that is dynamically modulated during embryonic development as well as during regenerative or pathological processes. Among the many model systems used in glycoscience research, we would like to propose the Xenopus laevis as an in vivo model system that is useful for corroborative studies aimed to answer specific, but still open questions regarding the role of extracellular matrix molecules, in dynamic cell behaviors such as proliferation, migration and differentiation. Our gene loss of function experiments demonstrated the requirement for the activity of UGDH in early phases of development and the role of HA in myogenesis by preventing early myoblasts from apoptosis. Moreover, we showed that CD44 is involved in hypaxial muscle cells migration. The evolutionary conservation of developmental mechanisms as well as gene functions leads to easier extrapolation of new “frog” findings to higher vertebrates thereby contributing to the further understanding of many aspects of cell biology.
Acknowledgements
These studies were supported by the Italian Ministry of University and Scientific Research (MIUR) and FAR. We thank Giancarlo De Luca, Manuela Viola, and Evgenia Karousou from the University of Insubria, and Irma Nardi and Paola Casini from the University of Pisa for reading and discussing the manuscript. A special thank goes to Roberto Perris (University of Parma).



