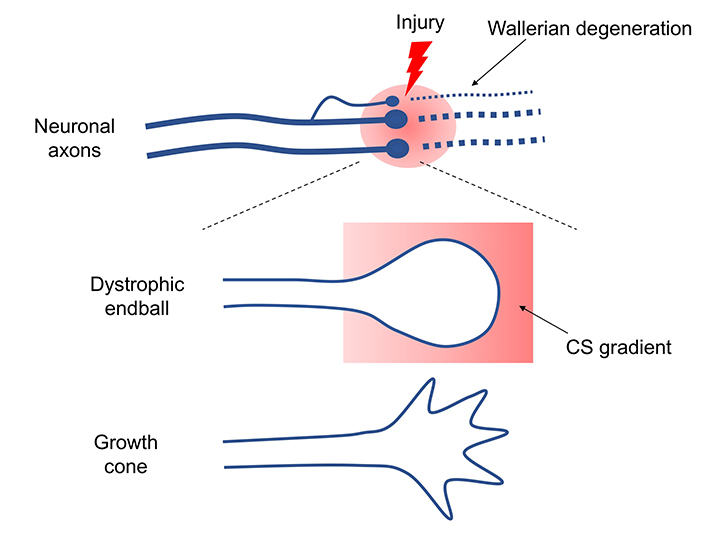
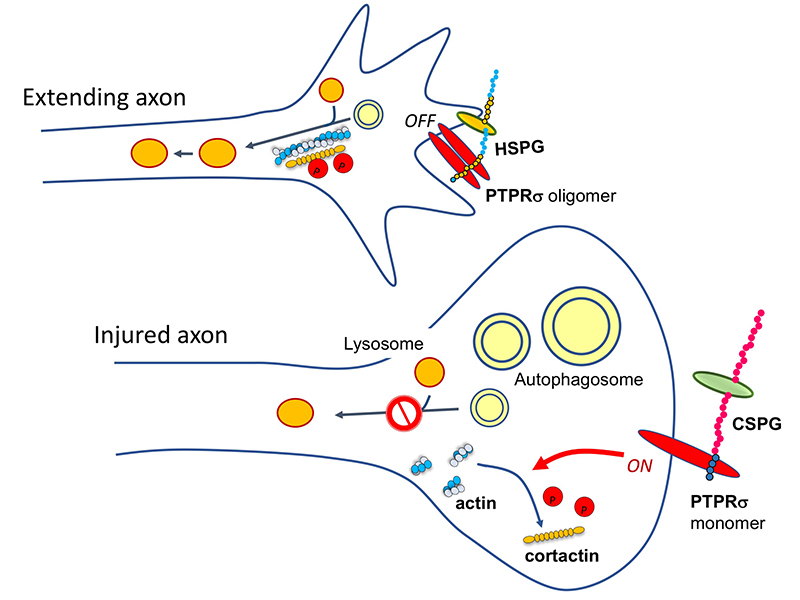
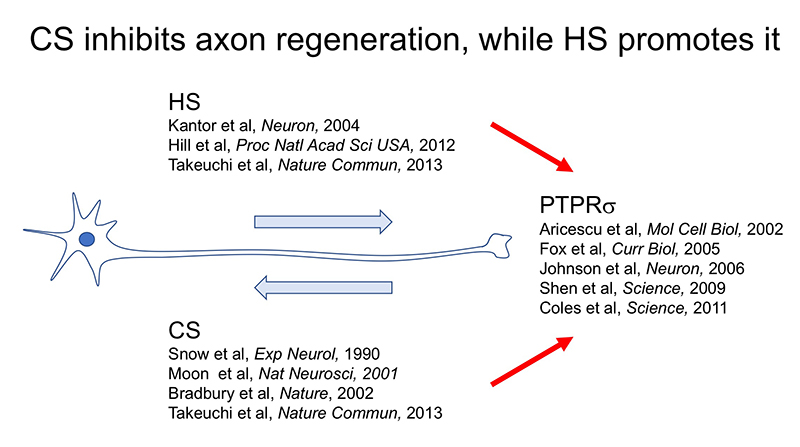
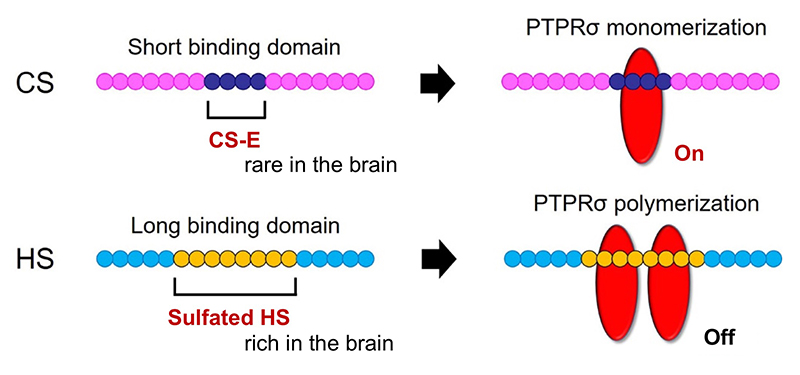
CSとPTPRσがもたらした軸索再生阻害機構はもう一つの観点からも重要である。それは軸索先端とオートファジーの連関である。正常なニューロンでは軸索の先端でオートファジーが始まり、autophagosome、autolysosomeが細胞体へ逆輸送されてオートファジー流が完成する(Wong, 2015)。従って軸索先端でこの流れが中断することによりdystrophic endball形成、軸索再生阻害が起きたという発見は、改めてニューロンにおけるオートファジーの重要性をハイライトしたことになる。この重要性は神経変性疾患にも適用できる。例えば家族性パーキンソン病でPINK1やParkinの遺伝子変異が見つかっているがこれはいずれもミトコンドリアのオートファジー(mitophagy)の重要なキナーゼとE3ユビキチンリガーゼである(Clark, 2006; Park, 2006; Yang, 2006)。このことと関連して、神経変性疾患ではニューロンの細胞死に先んじてシナプス変性が起こるというコンセプトが一般的に受け入れられている。神経変性のシナプス変性に伴ってautophagosomeの貯留がある。このような現象とCS-PTPRσの軸がどこまで重なるのか。興味深い視点の一つである。
References
Aricescu R, McKinnell IW, Halfter W, et al. Heparan sulfate proteoglycans are ligands for receptor protein tyrosine phosphatase σ. Mol Cell Biol. 22:1881–1892, 2002.
Bradbury EJ, Moon LD, Popat RJ, et al. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature. 416:636-640, 2002.
Clark ES, Whigham AS, Yarbrough WG, et al. Cortactin is an essential regulator of matrix metalloproteinase secretion and extracellular matrix degradation in invadopodia. Cancer Res. 67:4227-4235, 2007.
Clark IE, Dodson MW, Jiang C, et al. Drosophila pink1 Is required for mitochondrial function and interacts genetically with parkin. Nature. 441:1162-1166, 2006.
Coles CH, Mitakidis N, Zhang P, et al. Structural basis for extracellular cis and trans RPTPσ signal competition in synaptogenesis. Nat Commun. 5:5209, 2014.
Coles CH, Shen Y, Tenney AP, et al. Proteoglycan-specific molecular switch for RPTPσ clustering and neuronal extension. Science. 332:484-488, 2011.
Fox AN , Zinn K. The heparan sulfate proteoglycan syndecan is an in vivo ligand for the Drosophila LAR receptor tyrosine phosphatase. Curr Biol. 15:1701-1711, 2005.
Hasegawa J, Iwamoto R, Otomo T, et al. Autophagosome–lysosome fusion in neurons requires INPP5E, a protein associated with Joubert syndrome. EMBO J. 35:1853–1867, 2016.
Hill JJ, Jin K, Mao XO, et al. Intracerebral chondroitinase ABC and heparan sulfate proteoglycan glypican improve outcome from chronic stroke in rats. Proc Natl Acad Sci U S A. 109:9155-9160, 2012.
Ito Z, Sakamoto K, Imagama S, et al. N-acetylglucosamine 6-O-sulfotransferase-1-deficient mice show better functional recovery after spinal cord injury. J Neurosci. 30:5937-5947, 2010.
Johnson KG , Tenney AP, Ghose A, et al. The HSPGs Syndecan and Dallylike bind the receptor phosphatase LAR and exert distinct effects on synaptic development. Neuron. 49:517-531, 2006.
Kadomatsu K, Sakamoto K. Mechanisms of axon regeneration and its inhibition: roles of sulfated glycans. Arch Biochem Biophys. 558:36-41, 2014.
Kantor DB, Chivatakarn O, Peer KL, et al. Semaphorin 5A is a bifunctional axon guidance cue regulated by heparan and chondroitin sulfate proteoglycans. Neuron. 44:961-975, 2004.
Lang BT, Cregg JM, DePaul MA, et al. Modulation of the proteoglycan receptor PTPσ promotes recovery after spinal cord injury. Nature. 518:404-408, 2015.
Moon LD, Asher RA, Rhodes KE, et al. Regeneration of CNS axons back to their target following treatment of adult rat brain with chondroitinase ABC. Nat Neurosci. 4:465-466, 2001.
Naito Y, Lee AK, Takahashi H. Emerging roles of the neurotrophin receptor TrkC in synapse organization. Neurosci. Res. 116:10-17, 2017.
Park J, Lee SB, Lee S, et al. Mitochondrial dysfunction in Drosophila PINK1 mutants is complemented by parkin. Nature. 441:1157-1161, 2006.
Ruschel J, Hellal F, Flynn KC, et al. Systemic administration of epothilone B promotes axon regeneration after spinal cord injury. Science. 348:347-352, 2015.
Sakamoto K, Kadomatsu K. Mechanisms of axon regeneration: The significance of proteoglycans. Biochim Biophys Acta Gen Subj. 1861:2435-2441, 2017.
Sakamoto K, Ozaki T, Ko YC, et al. Glycan sulfation patterns define autophagy flux at axon tip via PTPRσ-cortactin axis. Nat Chem Biol. 15:699-709, 2019.
Shen Y, Tenney AP, Busch SA, et al. PTPsigma is a receptor for chondroitin sulfate proteoglycan, an inhibitor of neural regeneration. Science. 326:592-596, 2009.
Snow DM, Lemmon V, Carrino DA, et al. Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth in vitro. Exp Neurol. 109:111-130, 1990.
Takeuchi K, Yoshioka N, Onaga SH, et al. Chondroitin sulphate N-acetylgalactosaminyl-transferase-1 inhibits recovery from neural injury. Nat Commun. 4:2740, 2013.
Tom VJ, Steinmetz MP, Miller JH, et al. Studies on the development and behavior of the dystrophic growth cone, the Hallmark of regeneration failure, in an in vitro model of the glial scar and after spinal cord injury J. Neurosci. 24:6531-6539, 2004.
Tran AP, Warren PM, Silver J. Regulation of autophagy by inhibitory CSPG interactions with receptor PTPs and its impact on plasticity and regeneration after spinal cord injury. Exp Neurol. 328:113276, 2020.
Warren PM, Steiger SC, Dick TE, et al. Rapid and robust restoration of breathing long after spinal cord injury. Nat Commun. 9:4843, 2018.
Wong YC, Holzbaur ELF. Autophagosome dynamics in neurodegeneration at a glance. Journal of Cell Science. 128:1259-1267, 2015.
Yang Y, Gehrke S, Imai Y, et al. Mitochondrial pathology and muscle and dopaminergic neuron degeneration caused by inactivation of Drosophila Pink1 is rescued by Parkin. Proc Natl Acad Sci. 103:10793-10798, 2006.