編集者注1:英語の格言(The devil is in the details)で、「神は細部に宿る(英: God is in the details)」の転用とされる。細部に隠された罠や不可解な要素を見落としてはならないとの暗示で、一般に、一見、簡単にできそうなことでも、完成までに予想以上の労力を要すること、または、細部に隠されているために見つかりにくい重要な点に気が付かなかったために、失敗または異なる方向に進んでしまうことの意で用いられる。
ガレクチンと細胞表面糖タンパク質受容体間の架橋格子(lattice)形成モデル(図 1、表面に凝集しているガレクチン)は、いくつかの受容体のエンドサイトーシス阻害と、その阻害により増加した受容体によるシグナル伝達を説明するのに役立つ49,50。さらには、このような現象がGlcNAcの効果及びGlcNAcの代謝的アベイラビリティと関係するN型糖鎖の分岐のレベルと、どのように連動するのかを51を説明するためにも有益なモデルとなっている。ただし、最近のレビューで述べられているように、これに関する研究のほとんどは、細胞表面のガレクチン格子と、それにより影響を受ける糖タンパク質受容体のエンドサイトーシスに焦点を当てたものであり、ガレクチンそのもののエンドサイトーシスに注目したものではない52。他の研究では、上述のように、細胞に添加されたガレクチンは速やかにエンドサイトーシスされる傾向があり、さらにガレクチンにより、主にクラスリン非依存性エンドサイトーシスによる、特定の糖タンパク質のエンドサイトーシスが増加している可能性がある。ガレクチン-3についてはその詳細なメカニズムが提唱され、GlycoLipid-Lectin(GL-Lect)仮説と呼ばれている9,26。細胞表面に常在するガレクチン格子に関する証拠は十分とは言えないが、これは細胞の種類ごとに異なる可能性がある。かつて、とても入念にデザインされた研究手法において、細胞上のガレクチン-3凝集体がFRET(編集者注2)により可視化された53。それらの安定性の推定も試みられたと推察するが、細胞表面のガレクチンやエンドサイトーシス後の早期ガレクチンを識別することは困難であったと思われる。好中球では、凝集体が約10分間(無刺激細胞)、またはそれより短時間(刺激細胞)存続した。内皮細胞では、凝集体は細胞間結合部に認められ、蛍光ブリーチの30分後でも内在性ガレクチンの再分布が認められなかったことから、より安定しているように見えた。したがって、ほとんどの場合、ガレクチン相互作用は、架橋格子領域の分解と再編成により、速やかなターンオーバーをしているのではないか。このような緩やかな動態的見方では(編集者注:ガレクチン架橋格子が固定的なものでなく、形成と内部への取り込み(エンドサイトーシス)を繰り返しているため)、細胞表面のガレクチンや一時的なガレクチン格子が、異なる受容体のエンドサイトーシスを阻害したり増強したりすることができるのだろう(編集者注3)50。さらに、上述のように、エンドサイトーシスは可逆平衡ではなく、一方向性のプロセスである。ガレクチンが再び表面に出るためには、ソーティングに影響を及ぼす可能性のある、異なる局所環境(pHなど)を持つ種々のエンドソームから成るリサイクリング経路を通過しなければならない2,54。このことは、例えば標識ガレクチン-8をヒト線維芽細胞様細胞に添加した実験で示された(HFL-1、Carlsson, M. and Leffler, H.、未発表)。ガレクチン-8は数分後に細胞内小胞に認められたが、約10分後には、細胞膜近傍部位に集積し始めた。リサイクリング能を欠損したニーマンピックC由来の細胞では、ガレクチン-8が細胞内小胞にとどまり、膜に沿った集積を示さなかったことから、上記の集積はリサイクリングによるものと考えられる。このような結果を見ると、新たな疑問が生じてくる。リサイクリングしたガレクチンは、細胞外のガレクチン(外部から添加されたもの、または細胞膜にすでに存在しているもの)と比較した時、細胞表面において異なるプールに入るのか。もしそうであれば、それら二つの異なるプールはどのくらいの速さで相互作用し、混合するのか。
編集者注2:Fluorescence Resonance Energy Transferの略称。異なる波長特性をもつ蛍光色素で標識した分子同士の間でエネルギーのやり取りが起こる現象を利用して、分子間相互作用を解析する目的などに用いられる。
Johannes, L., Jacob, R., and Leffler, H. (2018) Galectins at a glance. J Cell Sci131, jcs208884
Banfer, S., and Jacob, R. (2020) Galectins in Intra- and Extracellular Vesicles. Biomolecules10, 1232
Mellman, I. (1996) Endocytosis and molecular sorting. Annu Rev Cell Dev Biol12, 575-625
Doherty, G. J., and McMahon, H. T. (2009) Mechanisms of endocytosis. Annu Rev Biochem78, 857-902
Mettlen, M., Chen, P. H., Srinivasan, S., Danuser, G., and Schmid, S. L. (2018) Regulation of Clathrin-Mediated Endocytosis. Annu Rev Biochem87, 871-896
Haudek, K. C., Spronk, K. J., Voss, P. G., Patterson, R. J., Wang, J. L., and Arnoys, E. J. (2010) Dynamics of galectin-3 in the nucleus and cytoplasm. Biochim Biophys Acta1800, 181-189
Kim, S. J., and Chun, K. H. (2020) Non-classical role of Galectin-3 in cancer progression: translocation to nucleus by carbohydrate-recognition independent manner. BMB Rep53, 173-180
Carlsson, M. C., Bengtson, P., Cucak, H., and Leffler, H. (2013) Galectin-3 guides intracellular trafficking of some human serotransferrin glycoforms. J Biol Chem288, 28398-28408
Lakshminarayan, R., Wunder, C., Becken, U., Howes, M. T., Benzing, C., Arumugam, S., Sales, S., Ariotti, N., Chambon, V., Lamaze, C., Loew, D., Shevchenko, A., Gaus, K., Parton, R. G., and Johannes, L. (2014) Galectin-3 drives glycosphingolipid-dependent biogenesis of clathrin-independent carriers. Nat Cell Biol16, 595-606
Renard, H. F., Tyckaert, F., Lo Giudice, C., Hirsch, T., Valades-Cruz, C. A., Lemaigre, C., Shafaq-Zadah, M., Wunder, C., Wattiez, R., Johannes, L., van der Bruggen, P., Alsteens, D., and Morsomme, P. (2020) Endophilin-A3 and Galectin-8 control the clathrin-independent endocytosis of CD166. Nat Commun11, 1457
Lindstedt, R., Apodaca, G., Barondes, S. H., Mostov, K. E., and Leffler, H. (1993) Apical secretion of a cytosolic protein by Madin-Darby canine kidney cells. Evidence for polarized release of an endogenous lectin by a nonclassical secretory pathway. J Biol Chem268, 11750-11757
Lingblom, C., Andersson, K., and Wenneras, C. (2021) Kinetic studies of galectin-10 release from eosinophils exposed to proliferating T cells. Clin Exp Immunol203, 230-243
Su, J. (2018) A Brief History of Charcot-Leyden Crystal Protein/Galectin-10 Research. Molecules23, 2931
Friedrichs, J., Torkko, J. M., Helenius, J., Teravainen, T. P., Fullekrug, J., Muller, D. J., Simons, K., and Manninen, A. (2007) Contributions of galectin-3 and -9 to epithelial cell adhesion analyzed by single cell force spectroscopy. J Biol Chem282, 29375-29383
Mishra, R., Grzybek, M., Niki, T., Hirashima, M., and Simons, K. (2010) Galectin-9 trafficking regulates apical-basal polarity in Madin-Darby canine kidney epithelial cells. Proc Natl Acad Sci U S A107, 17633-17638
Elia, A. E., Boardman, A. P., Wang, D. C., Huttlin, E. L., Everley, R. A., Dephoure, N., Zhou, C., Koren, I., Gygi, S. P., and Elledge, S. J. (2015) Quantitative Proteomic Atlas of Ubiquitination and Acetylation in the DNA Damage Response. Mol Cell59, 867-881
Sato, S., and Hughes, R. C. (1994) Regulation of secretion and surface expression of Mac-2, a galactoside-binding protein of macrophages. J Biol Chem269, 4424-4430
Beyer, E. C., and Barondes, S. H. (1982) Secretion of endogenous lectin by chicken intestinal goblet cells. J Cell Biol92, 28-33
Bhat, R., Belardi, B., Mori, H., Kuo, P., Tam, A., Hines, W. C., Le, Q. T., Bertozzi, C. R., and Bissell, M. J. (2016) Nuclear repartitioning of galectin-1 by an extracellular glycan switch regulates mammary morphogenesis. Proc Natl Acad Sci U S A113, E4820-4827
Seelenmeyer, C., Wegehingel, S., Tews, I., Kunzler, M., Aebi, M., and Nickel, W. (2005) Cell surface counter receptors are essential components of the unconventional export machinery of galectin-1. J Cell Biol171, 373-381
Ochieng, J., Green, B., Evans, S., James, O., and Warfield, P. (1998) Modulation of the biological functions of galectin-3 by matrix metalloproteinases. Biochim Biophys Acta1379, 97-106
Nangia-Makker, P., Balan, V., and Raz, A. (2008) Regulation of tumor progression by extracellular galectin-3. Cancer Microenviron1, 43-51
Nieminen, J., St-Pierre, C., and Sato, S. (2005) Galectin-3 interacts with naive and primed neutrophils, inducing innate immune responses. J Leukoc Biol78, 1127-1135
Nishi, N., Itoh, A., Shoji, H., Miyanaka, H., and Nakamura, T. (2006) Galectin-8 and galectin-9 are novel substrates for thrombin. Glycobiology16, 15C-20C
Furtak, V., Hatcher, F., and Ochieng, J. (2001) Galectin-3 mediates the endocytosis of beta-1 integrins by breast carcinoma cells. Biochem Biophys Res Commun 289, 845-850
Johannes, L. (2021) The Cellular and Chemical Biology of Endocytic Trafficking and Intracellular Delivery-The GL-Lect Hypothesis. Molecules26, 3299
Paz, I., Sachse, M., Dupont, N., Mounier, J., Cederfur, C., Enninga, J., Leffler, H., Poirier, F., Prevost, M. C., Lafont, F., and Sansonetti, P. (2010) Galectin-3, a marker for vacuole lysis by invasive pathogens. Cell Microbiol12, 530-544
Jia, J., Claude-Taupin, A., Gu, Y., Choi, S. W., Peters, R., Bissa, B., Mudd, M. H., Allers, L., Pallikkuth, S., Lidke, K. A., Salemi, M., Phinney, B., Mari, M., Reggiori, F., and Deretic, V. (2020) Galectin-3 Coordinates a Cellular System for Lysosomal Repair and Removal. Dev Cell52, 69-87 e68
Jia, J., Claude-Taupin, A., Gu, Y., Choi, S. W., Peters, R., Bissa, B., Mudd, M. H., Allers, L., Pallikkuth, S., Lidke, K. A., Salemi, M., Phinney, B., Mari, M., Reggiori, F., and Deretic, V. (2020) MERIT, a cellular system coordinating lysosomal repair, removal and replacement. Autophagy16, 1539-1541
Lepur, A., Carlsson, M. C., Novak, R., Dumic, J., Nilsson, U. J., and Leffler, H. (2012) Galectin-3 endocytosis by carbohydrate independent and dependent pathways in different macrophage like cell types. Biochim Biophys Acta1820, 804-818
Horie, H., and Kadoya, T. (2002) Galectin-1 plays essential roles in adult mammalian nervous tissues. Roles of oxidized galectin-1. Glycoconj J19, 479-489
Nishi, N., Abe, A., Iwaki, J., Yoshida, H., Itoh, A., Shoji, H., Kamitori, S., Hirabayashi, J., and Nakamura, T. (2008) Functional and structural bases of a cysteine-less mutant as a long-lasting substitute for galectin-1. Glycobiology18, 1065-1073
Karlsson, A., Follin, P., Leffler, H., and Dahlgren, C. (1998) Galectin-3 activates the NADPH-oxidase in exudated but not peripheral blood neutrophils. Blood91, 3430-3438
Stowell, S. R., Qian, Y., Karmakar, S., Koyama, N. S., Dias-Baruffi, M., Leffler, H., McEver, R. P., and Cummings, R. D. (2008) Differential roles of galectin-1 and galectin-3 in regulating leukocyte viability and cytokine secretion. J Immunol180, 3091-3102
Du Rietz, H., Hedlund, H., Wilhelmson, S., Nordenfelt, P., and Wittrup, A. (2020) Imaging small molecule-induced endosomal escape of siRNA. Nat Commun11, 1809
Stegmayr, J., Zetterberg, F., Carlsson, M. C., Huang, X., Sharma, G., Kahl-Knutson, B., Schambye, H., Nilsson, U. J., Oredsson, S., and Leffler, H. (2019) Extracellular and intracellular small-molecule galectin-3 inhibitors. Sci Rep9, 2186
Lepur, A., Salomonsson, E., Nilsson, U. J., and Leffler, H. (2012) Ligand induced galectin-3 protein self-association. J Biol Chem287, 21751-21756
Salomonsson, E., Larumbe, A., Tejler, J., Tullberg, E., Rydberg, H., Sundin, A., Khabut, A., Frejd, T., Lobsanov, Y. D., Rini, J. M., Nilsson, U. J., and Leffler, H. (2010) Monovalent interactions of galectin-1. Biochemistry49, 9518-9532
Leppanen, A., Stowell, S., Blixt, O., and Cummings, R. D. (2005) Dimeric galectin-1 binds with high affinity to alpha2,3-sialylated and non-sialylated terminal N-acetyllactosamine units on surface-bound extended glycans. J Biol Chem280, 5549-5562
Leppanen, A., Arthur, C. M., Stowell, S. R., and Cummings, R. D. (2022) Examination of Whole-Cell Galectin Binding by Solid Phase and Flow Cytometric Analysis. Methods Mol Biol2442, 187-203
Nielsen, M. I., Stegmayr, J., Grant, O. C., Yang, Z., Nilsson, U. J., Boos, I., Carlsson, M. C., Woods, R. J., Unverzagt, C., Leffler, H., and Wandall, H. H. (2018) Galectin binding to cells and glycoproteins with genetically modified glycosylation reveals galectin-glycan specificities in a natural context. J Biol Chem293, 20249-20262
Stowell, S. R., Arthur, C. M., Mehta, P., Slanina, K. A., Blixt, O., Leffler, H., Smith, D. F., and Cummings, R. D. (2008) Galectin-1, -2, and -3 exhibit differential recognition of sialylated glycans and blood group antigens. J Biol Chem283, 10109-10123
Carlsson, S., Oberg, C. T., Carlsson, M. C., Sundin, A., Nilsson, U. J., Smith, D., Cummings, R. D., Almkvist, J., Karlsson, A., and Leffler, H. (2007) Affinity of galectin-8 and its carbohydrate recognition domains for ligands in solution and at the cell surface. Glycobiology17, 663-676
Salomonsson, E., Carlsson, M. C., Osla, V., Hendus-Altenburger, R., Kahl-Knutson, B., Oberg, C. T., Sundin, A., Nilsson, R., Nordberg-Karlsson, E., Nilsson, U. J., Karlsson, A., Rini, J. M., and Leffler, H. (2010) Mutational tuning of galectin-3 specificity and biological function. J Biol Chem285, 35079-35091
Peterson, K., Kumar, R., Stenstrom, O., Verma, P., Verma, P. R., Hakansson, M., Kahl-Knutsson, B., Zetterberg, F., Leffler, H., Akke, M., Logan, D. T., and Nilsson, U. J. (2018) Systematic Tuning of Fluoro-galectin-3 Interactions Provides Thiodigalactoside Derivatives with Single-Digit nM Affinity and High Selectivity. J Med Chem61, 1164-1175
Zetterberg, F. R., Peterson, K., Johnsson, R. E., Brimert, T., Hakansson, M., Logan, D. T., Leffler, H., and Nilsson, U. J. (2018) Monosaccharide Derivatives with Low-Nanomolar Lectin Affinity and High Selectivity Based on Combined Fluorine-Amide, Phenyl-Arginine, Sulfur-pi, and Halogen Bond Interactions. ChemMedChem13, 133-137
Perillo, N. L., Pace, K. E., Seilhamer, J. J., and Baum, L. G. (1995) Apoptosis of T cells mediated by galectin-1. Nature378, 736-739
Stillman, B. N., Hsu, D. K., Pang, M., Brewer, C. F., Johnson, P., Liu, F. T., and Baum, L. G. (2006) Galectin-3 and galectin-1 bind distinct cell surface glycoprotein receptors to induce T cell death. J Immunol176, 778-789
Nabi, I. R., Shankar, J., and Dennis, J. W. (2015) The galectin lattice at a glance. J Cell Sci128, 2213-2219
Mathew, M. P., and Donaldson, J. G. (2018) Distinct cargo-specific response landscapes underpin the complex and nuanced role of galectin-glycan interactions in clathrin-independent endocytosis. J Biol Chem293, 7222-7237
Lau, K. S., Partridge, E. A., Grigorian, A., Silvescu, C. I., Reinhold, V. N., Demetriou, M., and Dennis, J. W. (2007) Complex N-glycan number and degree of branching cooperate to regulate cell proliferation and differentiation. Cell129, 123-134
Nieminen, J., Kuno, A., Hirabayashi, J., and Sato, S. (2007) Visualization of galectin-3 oligomerization on the surface of neutrophils and endothelial cells using fluorescence resonance energy transfer. J Biol Chem282, 1374-1383
von Mach, T., Carlsson, M. C., Straube, T., Nilsson, U., Leffler, H., and Jacob, R. (2014) Ligand binding and complex formation of galectin-3 is modulated by pH variations. Biochem J457, 107-115
Iwaki, J., and Hirabayashi, J. (2015) Evaluation of galectin binding by frontal affinity chromatography (FAC). Methods Mol Biol 1207, 63-74
Leffler, H., and Barondes, S. H. (1986) Specificity of binding of three soluble rat lung lectins to substituted and unsubstituted mammalian beta-galactosides. J Biol Chem261, 10119-10126
Varki, A., Cummings, R. D., Esko, J. D., Stanley, P., Hart, G. W., Aebi, M., Mohnen, D., Kinoshita, T., Packer, N. H., Prestegard, J. H., Schnaar, R. L., and Seeberger, P. H. (2022) Essentials of glycobiology, Fourth edition. ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Patnaik, S. K., Potvin, B., Carlsson, S., Sturm, D., Leffler, H., and Stanley, P. (2006) Complex N-glycans are the major ligands for galectin-1, -3, and -8 on Chinese hamster ovary cells. Glycobiology16, 305-317
Sato, S., and Hughes, R. C. (1992) Binding specificity of a baby hamster kidney lectin for H type I and II chains, polylactosamine glycans, and appropriately glycosylated forms of laminin and fibronectin. J Biol Chem267, 6983-6990
Gao, C., Hanes, M. S., Byrd-Leotis, L. A., Wei, M., Jia, N., Kardish, R. J., McKitrick, T. R., Steinhauer, D. A., and Cummings, R. D. (2019) Unique Binding Specificities of Proteins toward Isomeric Asparagine-Linked Glycans. Cell Chem Biol26, 535-547 e534
Carlsson, M. C., Cederfur, C., Schaar, V., Balog, C. I., Lepur, A., Touret, F., Salomonsson, E., Deelder, A. M., Ferno, M., Olsson, H., Wuhrer, M., and Leffler, H. (2011) Galectin-1-binding glycoforms of haptoglobin with altered intracellular trafficking, and increase in metastatic breast cancer patients. PLoS One6, e26560
St-Pierre, C., Manya, H., Ouellet, M., Clark, G. F., Endo, T., Tremblay, M. J., and Sato, S. (2011) Host-soluble galectin-1 promotes HIV-1 replication through a direct interaction with glycans of viral gp120 and host CD4. J Virol85, 11742-11751
Mirgorodskaya, E., Dransart, E., Shafaq-Zadah, M., Roderer, D., Sihlbom, C., Leffler, H., and Johannes, L. (2022) Site-specific N-glycan profiles of alpha5 beta1 integrin from rat liver. Biol Cell114, 160-176
Carlsson, S., Carlsson, M. C., and Leffler, H. (2007) Intracellular sorting of galectin-8 based on carbohydrate fine specificity. Glycobiology17, 906-912
Ideo, H., Seko, A., Ishizuka, I., and Yamashita, K. (2003) The N-terminal carbohydrate recognition domain of galectin-8 recognizes specific glycosphingolipids with high affinity. Glycobiology13, 713-723
Hong, M. H., Lin, W. H., Weng, I. C., Hung, Y. H., Chen, H. L., Chen, H. Y., Chen, P., Lin, C. H., Yang, W. Y., and Liu, F. T. (2019) Intracellular galectins control cellular responses commensurate with cell surface carbohydrate composition. Glycobiology30, 49-57
Blenda, A. V., Kamili, N. A., Wu, S. C., Abel, W. F., Ayona, D., Gerner-Smidt, C., Ho, A. D., Benian, G. M., Cummings, R. D., Arthur, C. M., and Stowell, S. R. (2022) Galectin-9 recognizes and exhibits antimicrobial activity toward microbes expressing blood group-like antigens. J Biol Chem298, 101704
Lukyanov, P., Furtak, V., and Ochieng, J. (2005) Galectin-3 interacts with membrane lipids and penetrates the lipid bilayer. Biochem Biophys Res Commun338, 1031-1036
Dennis, J. W. (2015) Many Light Touches Convey the Message. Trends Biochem Sci40, 673-686
Lin, Y. H., Qiu, D. C., Chang, W. H., Yeh, Y. Q., Jeng, U. S., Liu, F. T., and Huang, J. R. (2017) The intrinsically disordered N-terminal domain of galectin-3 dynamically mediates multisite self-association of the protein through fuzzy interactions. J Biol Chem292, 17845-17856
Chiu, Y. P., Sun, Y. C., Qiu, D. C., Lin, Y. H., Chen, Y. Q., Kuo, J. C., and Huang, J. R. (2020) Liquid-liquid phase separation and extracellular multivalent interactions in the tale of galectin-3. Nat Commun11, 1229
Daussy, C. F., Pied, N., and Wodrich, H. (2021) Understanding Post Entry Sorting of Adenovirus Capsids; A Chance to Change Vaccine Vector Properties. Viruses13, 1221
Hong, M. H., Weng, I. C., Li, F. Y., Lin, W. H., and Liu, F. T. (2021) Intracellular galectins sense cytosolically exposed glycans as danger and mediate cellular responses. J Biomed Sci28, 16
Vuong, L., Kouverianou, E., Rooney, C. M., McHugh, B. J., Howie, S. E. M., Gregory, C. D., Forbes, S. J., Henderson, N. C., Zetterberg, F. R., Nilsson, U. J., Leffler, H., Ford, P., Pedersen, A., Gravelle, L., Tantawi, S., Schambye, H., Sethi, T., and MacKinnon, A. C. (2019) An Orally Active Galectin-3 Antagonist Inhibits Lung Adenocarcinoma Growth and Augments Response to PD-L1 Blockade. Cancer Res79, 1480-1492
Slack, R. J., Mills, R., and Mackinnon, A. C. (2021) The therapeutic potential of galectin-3 inhibition in fibrotic disease. Int J Biochem Cell Biol130, 105881
Croci, D. O., Cerliani, J. P., Dalotto-Moreno, T., Mendez-Huergo, S. P., Mascanfroni, I. D., Dergan-Dylon, S., Toscano, M. A., Caramelo, J. J., Garcia-Vallejo, J. J., Ouyang, J., Mesri, E. A., Junttila, M. R., Bais, C., Shipp, M. A., Salatino, M., and Rabinovich, G. A. (2014) Glycosylation-dependent lectin-receptor interactions preserve angiogenesis in anti-VEGF refractory tumors. Cell156, 744-758
Hirani, N., MacKinnon, A. C., Nicol, L., Ford, P., Schambye, H., Pedersen, A., Nilsson, U. J., Leffler, H., Sethi, T., Tantawi, S., Gravelle, L., Slack, R. J., Mills, R., Karmakar, U., Humphries, D., Zetterberg, F., Keeling, L., Paul, L., Molyneaux, P. L., Li, F., Funston, W., Forrest, I. A., Simpson, A. J., Gibbons, M. A., and Maher, T. M. (2021) Target inhibition of galectin-3 by inhaled TD139 in patients with idiopathic pulmonary fibrosis. Eur Respir J57, 2002559
Kim, S. J., Kang, H. G., Kim, K., Kim, H., Zetterberg, F., Park, Y. S., Cho, H. S., Hewitt, S. M., Chung, J. Y., Nilsson, U. J., Leffler, H., and Chun, K. H. (2021) Crosstalk between WNT and STAT3 is mediated by galectin-3 in tumor progression. Gastric Cancer24, 1050-1062
Stegmayr, J., Lepur, A., Kahl-Knutson, B., Aguilar-Moncayo, M., Klyosov, A. A., Field, R. A., Oredsson, S., Nilsson, U. J., and Leffler, H. (2016) Low or No Inhibitory Potency of the Canonical Galectin Carbohydrate-binding Site by Pectins and Galactomannans. J Biol Chem291, 13318-13334
Pedrosa, L. F., Raz, A., and Fabi, J. P. (2022) The Complex Biological Effects of Pectin: Galectin-3 Targeting as Potential Human Health Improvement? Biomolecules12, 289

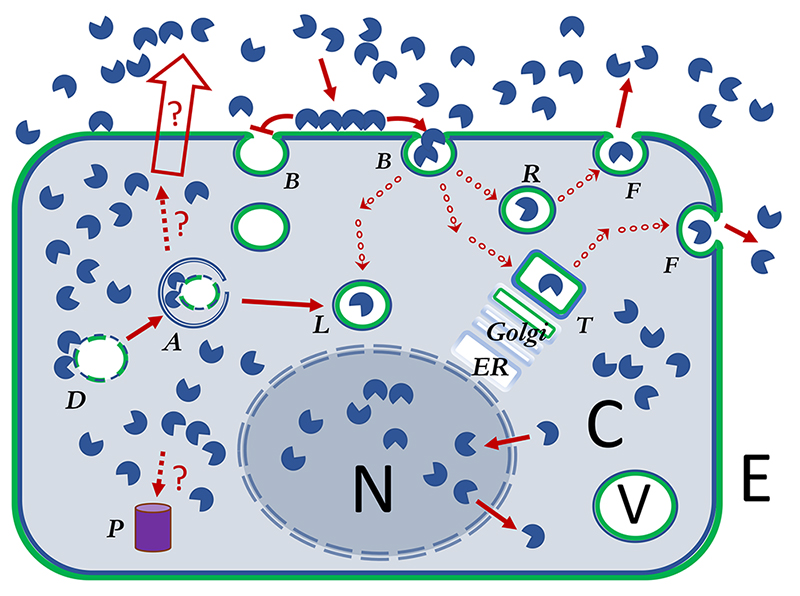
 すべてのタイプのガレクチンは一量体CRDとして表している。
すべてのタイプのガレクチンは一量体CRDとして表している。 ガレクチンによる細胞表面リガンドの架橋(cross-linking)は細胞表面の糖タンパク質受容体発現を制御していると提唱されている。すなわち、エンドサイトーシスによる受容体の取り込みを阻害(ガレクチン格子(lattice)仮説)、あるいは、促進(GL-Lect仮説)する。
ガレクチンによる細胞表面リガンドの架橋(cross-linking)は細胞表面の糖タンパク質受容体発現を制御していると提唱されている。すなわち、エンドサイトーシスによる受容体の取り込みを阻害(ガレクチン格子(lattice)仮説)、あるいは、促進(GL-Lect仮説)する。 濃い赤の矢印は細胞内外でのガレクチンの経路または影響を示している。本論文と関連する未解明の経路は、点線の矢印と?マークで示されている。
濃い赤の矢印は細胞内外でのガレクチンの経路または影響を示している。本論文と関連する未解明の経路は、点線の矢印と?マークで示されている。 小さな丸の連なった矢印は、ガレクチンの細胞内輸送経路を示す。小胞(~100 nm)に取り込まれたガレクチンは、リソソーム、トランスゴルジネットワーク、様々なタイプのエンドソームやエクソソームといった特定機能を持つ様々な比較的大きな小胞(~1000nm)へ輸送される。
小さな丸の連なった矢印は、ガレクチンの細胞内輸送経路を示す。小胞(~100 nm)に取り込まれたガレクチンは、リソソーム、トランスゴルジネットワーク、様々なタイプのエンドソームやエクソソームといった特定機能を持つ様々な比較的大きな小胞(~1000nm)へ輸送される。
 塗りつぶしなしの矢印は、まだメカニズムが不明確の非古典的な(non-classical)ガレクチン分泌を示している。
塗りつぶしなしの矢印は、まだメカニズムが不明確の非古典的な(non-classical)ガレクチン分泌を示している。
 )または、阻害(
)または、阻害( )されるエンドサイトーシスによる膜陥入、ピットの形成及び細胞内へのブッディング
)されるエンドサイトーシスによる膜陥入、ピットの形成及び細胞内へのブッディング


