Cipollo JF, Parsons LM. Glycomics and glycoproteomics of viruses: Mass spectrometry applications and insights toward structure-function relationships. Mass Spectrom Rev. 2020 Jul;39(4):371-409.
Vreeker GCM, Vangangelt KMH, Bladergroen MR, Nicolardi S, Mesker WE, Wuhrer M, van der Burgt YEM, Tollenaar RAEM. Serum N-glycan profiles differ for various breast cancer subtypes. Glycoconj J. 2021 Jun;38(3):387-395.
Miura Y, Arai T, Yamagata T. Synthesis of amphiphilic lactosides that possess a lactosylceramide-mimicking N-acyl structure: alternative universal substrates for endo-type glycosylceramidases. Carbohydr Res. 1996 Aug 19;289:193-9.
Miura Y, Yamagata T. Glycosylation of lactosylceramide analogs in animal cells: amphipathic disaccharide primers for glycosphingolipid synthesis. Biochem Biophys Res Commun. 1997 Dec 29;241(3):698-703.
Nakajima H, Miura Y, Yamagata T. Glycosylation of amphipathic lactoside primers with consequent inhibition of endogenous glycosphingolipid synthesis. J Biochem. 1998 Jul;124(1):148-56.
Sato T, Takashiba M, Hayashi R, Zhu X, Yamagata T. Glycosylation of dodecyl 2-acetamido-2-deoxy-beta-D-glucopyranoside and dodecyl beta-D-galactopyranosyl-(1-->4)-2-acetamido-2-deoxy-beta-D-glucopyranoside as saccharide primers in cells. Carbohydr Res. 2008 Apr 7;343(5):831-8.
Sakura R, Nagai K, Yagi Y, Takahashi Y, Ide Y, Yagi Y, Yamamoto D, Mizuno M, Sato T. In vitro synthesis of mucin-type O-glycans using saccharide primers comprising GalNAc-Ser and GalNAc-Thr residues. Carbohydr Res. 2022 Jan;511:108495.
Wang Y, Kumazawa T, Shiba K, Osumi K, Mizuno M, Sato T. Glycosylation of Nα-lauryl-O-(β-D-xylopyranosyl)-L-serinamide as a saccharide primer in cells. Carbohydr Res. 2012 Nov 1;361:33-40.
Wang Y, Yang X, Yamagata S, Yamagata T, Sato T. Involvement of Ext1 and heparanase in migration of mouse FBJ osteosarcoma cells. Mol Cell Biochem. 2013 Jan;373(1-2):63-72.
Otsuka Y, Sato T. Comparative Quantification Method for Glycosylated Products Elongated on β-Xylosides Using a Stable Isotope-Labeled Saccharide Primer. Anal Chem. 2018 Apr 17;90(8):5201-5208.
Otsuka Y, Sato T. Saccharide Primers Comprising Xylosyl-Serine Primed Phosphorylated Oligosaccharides Act as Intermediates in Glycosaminoglycan Biosynthesis. ACS Omega. 2017 Jul 31;2(7):3110-3122.
Stein MN, Patel N, Bershadskiy A, Sokoloff A, Singer EA. Androgen synthesis inhibitors in the treatment of castration-resistant prostate cancer. Asian J Androl. 2014 May-Jun;16(3):387-400.
Castro-Vázquez G. Cultural Scripts Underpinning Prostate Cancer-Literacy in Japan. Am J Mens Health. 2022 Jan-Feb;16(1):15579883221076658.
Van Poppel H, Klotz L. Gonadotropin-releasing hormone: an update review of the antagonists versus agonists. Int J Urol. 2012 Jul;19(7):594-601.
Shigeta K, Kosaka T, Hongo H, Yanai Y, Matsumoto K, Morita S, Mizuno R, Shinojima T, Kikuchi E, Oya M. Castration-resistant prostate cancer patients who had poor response on first androgen deprivation therapy would obtain certain clinical benefit from early docetaxel administration. Int J Clin Oncol. 2019 May;24(5):546-553.
Rodriguez-Vida A, Galazi M, Rudman S, Chowdhury S, Sternberg CN. Enzalutamide for the treatment of metastatic castration-resistant prostate cancer. Drug Des Devel Ther. 2015 Jun 29;9:3325-39.
Balk SP, Ko YJ, Bubley GJ. Biology of prostate-specific antigen. J Clin Oncol. 2003 Jan 15;21(2):383-91.
De Palma M, Biziato D, Petrova TV. Microenvironmental regulation of tumour angiogenesis. Nat Rev Cancer. 2017 Aug;17(8):457-474.
Lim B, Woodward WA, Wang X, Reuben JM, Ueno NT. Inflammatory breast cancer biology: the tumour microenvironment is key. Nat Rev Cancer. 2018 Aug;18(8):485-499.
Greville G, Llop E, Huang C, Creagh-Flynn J, Pfister S, O'Flaherty R, Madden SF, Peracaula R, Rudd PM, McCann A, Saldova R. Hypoxia Alters Epigenetic and N-Glycosylation Profiles of Ovarian and Breast Cancer Cell Lines in-vitro. Front Oncol. 2020 Jul 29;10:1218.
Arriagada C, Silva P, Torres VA. Role of glycosylation in hypoxia-driven cell migration and invasion. Cell Adh Migr. 2019 Dec;13(1):13-22.
Ma S, Zhao Y, Lee WC, Ong LT, Lee PL, Jiang Z, Oguz G, Niu Z, Liu M, Goh JY, Wang W, Bustos MA, Ehmsen S, Ramasamy A, Hoon DSB, Ditzel HJ, Tan EY, Chen Q, Yu Q. Hypoxia induces HIF1α-dependent epigenetic vulnerability in triple negative breast cancer to confer immune effector dysfunction and resistance to anti-PD-1 immunotherapy. Nat Commun. 2022 Jul 15;13(1):4118.
Hochachka PW, Rupert JL, Goldenberg L, Gleave M, Kozlowski P. Going malignant: the hypoxia-cancer connection in the prostate. Bioessays. 2002 Aug;24(8):749-57.
Geng H, Xue C, Mendonca J, Sun XX, Liu Q, Reardon PN, Chen Y, Qian K, Hua V, Chen A, Pan F, Yuan J, Dang S, Beer TM, Dai MS, Kachhap SK, Qian DZ. Interplay between hypoxia and androgen controls a metabolic switch conferring resistance to androgen/AR-targeted therapy. Nat Commun. 2018 Nov 26;9(1):4972.
Fraga A, Ribeiro R, Príncipe P, Lopes C, Medeiros R. Hypoxia and Prostate Cancer Aggressiveness: A Tale With Many Endings. Clin Genitourin Cancer. 2015 Aug;13(4):295-301.
Tan Z, Wang C, Li X, Guan F. Bisecting N-Acetylglucosamine Structures Inhibit Hypoxia-Induced Epithelial-Mesenchymal Transition in Breast Cancer Cells. Front Physiol. 2018 Mar 9;9:210.
Otsuki H, Kimura T, Yamaga T, Kosaka T, Suehiro JI, Sakurai H. Prostate Cancer Cells in Different Androgen Receptor Status Employ Different Leucine Transporters. Prostate. 2017 Feb;77(2):222-233.
Cervoni GE, Cheng JJ, Stackhouse KA, Heimburg-Molinaro J, Cummings RD. O-glycan recognition and function in mice and human cancers. Biochem J. 2020 Apr 30;477(8):1541-1564.
Sakuma K, Aoki M, Kannagi R. Transcription factors c-Myc and CDX2 mediate E-selectin ligand expression in colon cancer cells undergoing EGF/bFGF-induced epithelial-mesenchymal transition. Proc Natl Acad Sci U S A. 2012 May 15;109(20):7776-81.
Cicchini C, de Nonno V, Battistelli C, Cozzolino AM, De Santis Puzzonia M, Ciafrè SA, Brocker C, Gonzalez FJ, Amicone L, Tripodi M. Epigenetic control of EMT/MET dynamics: HNF4α impacts DNMT3s through miRs-29. Biochim Biophys Acta. 2015 Aug;1849(8):919-29.
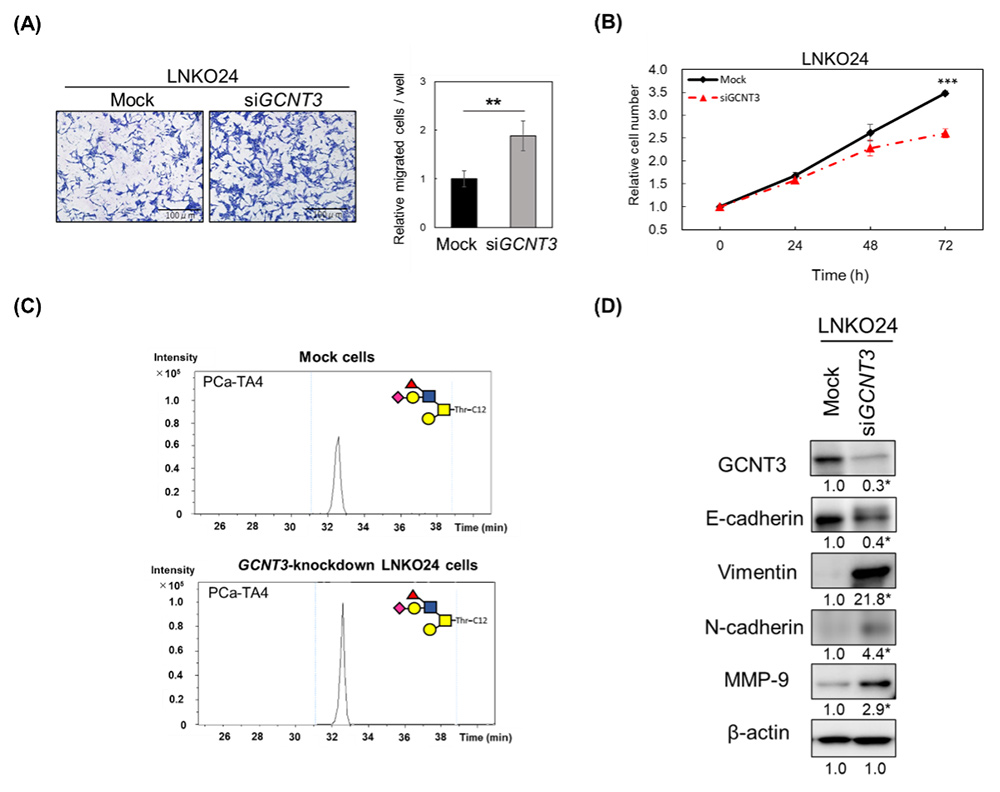
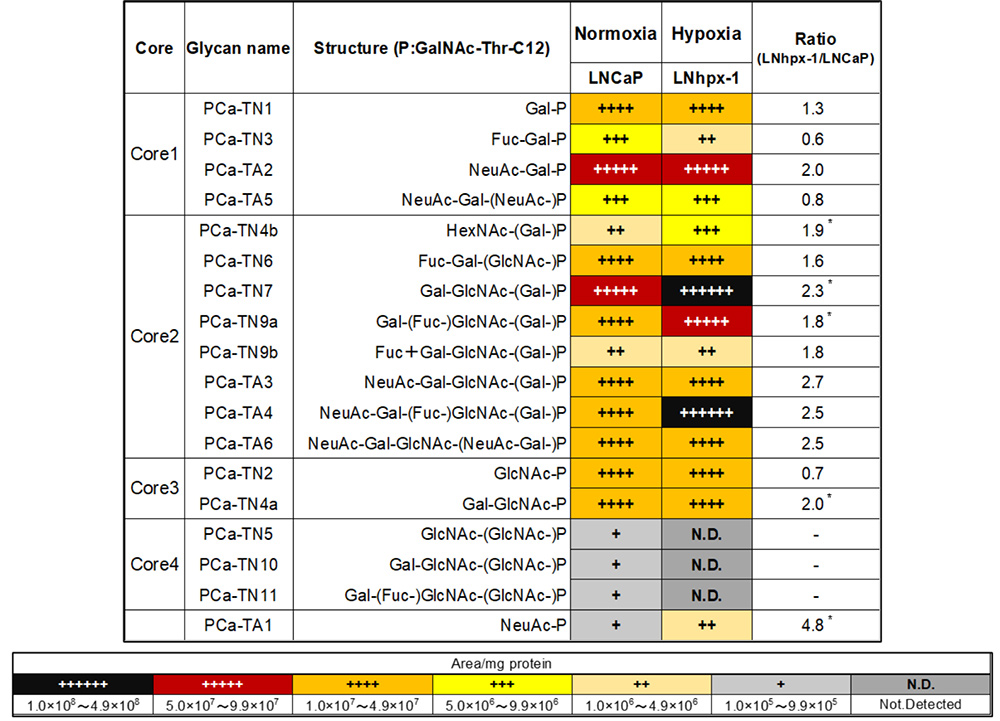
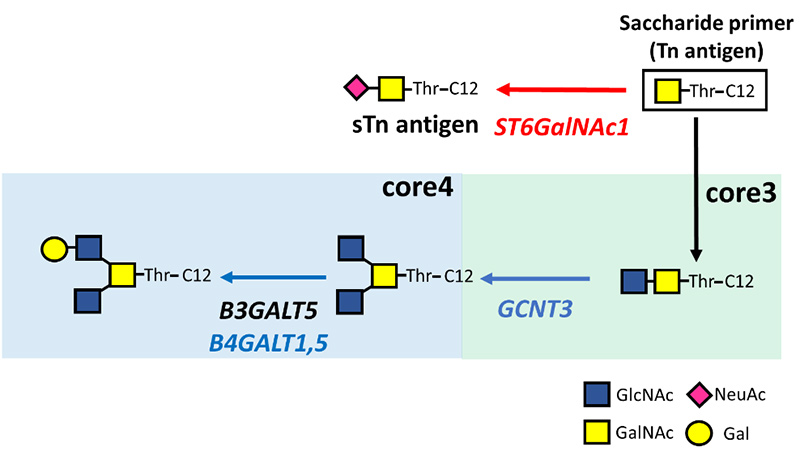
Yamamoto D, Sasaki K, Kosaka T, Oya M, Sato T. Functional analysis of GCNT3 for cell migration and EMT of castration-resistant prostate cancer cells. Glycobiology. 2022 Sep 19;32(10):897-908.
Zhang Y, Hong Y, Wang D, Duan L, Liu Y, Li L, Liu D, Zhuang K, Wei C, Zheng G, Huo C, Liu G. Hsa_circ_0076305 induces migration-proliferation dichotomy in gastric cancer. Oncol Lett. 2021 Mar;21(3):220.
Bhandari D, Lopez-Sanchez I, To A, Lo IC, Aznar N, Leyme A, Gupta V, Niesman I, Maddox AL, Garcia-Marcos M, Farquhar MG, Ghosh P. Cyclin-dependent kinase 5 activates guanine nucleotide exchange factor GIV/Girdin to orchestrate migration-proliferation dichotomy. Proc Natl Acad Sci U S A. 2015 Sep 1;112(35):E4874-83.
Chen C, Enomoto A, Weng L, Taki T, Shiraki Y, Mii S, Ichihara R, Kanda M, Koike M, Kodera Y, Takahashi M. Complex roles of the actin-binding protein Girdin/GIV in DNA damage-induced apoptosis of cancer cells. Cancer Sci. 2020 Nov;111(11):4303-4317.
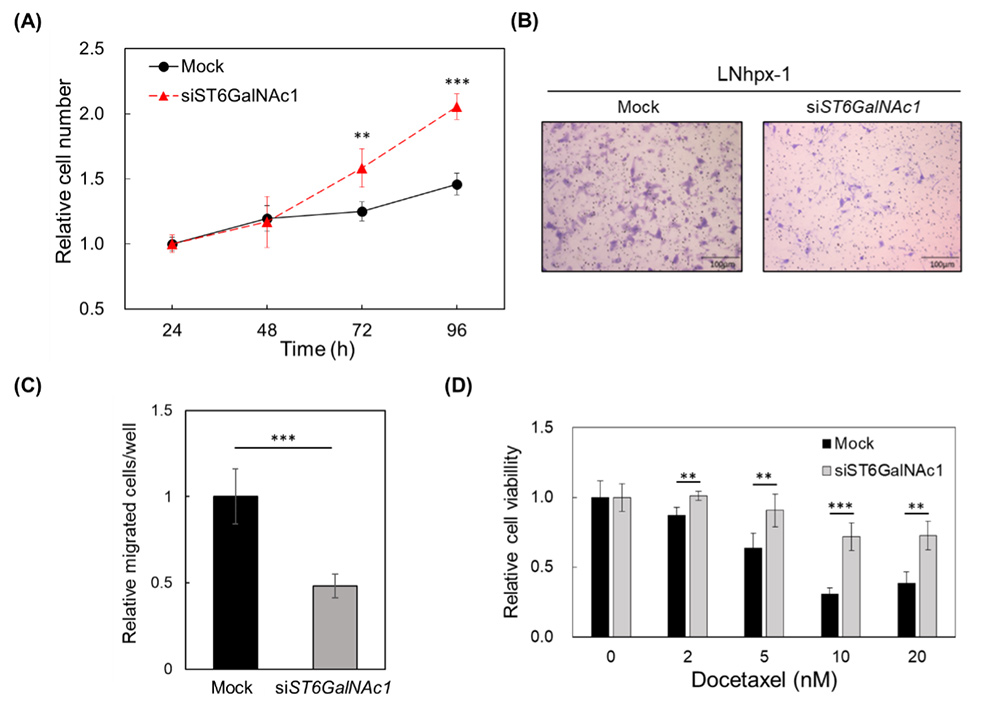
Yamamoto D, Hongo H, Kosaka T, Aoki N, Oya M, Sato T. The sialyl-Tn antigen synthase genes regulates migration-proliferation dichotomy in prostate cancer cells under hypoxia. Glycoconj J. 2023 Apr;40(2):199-212.