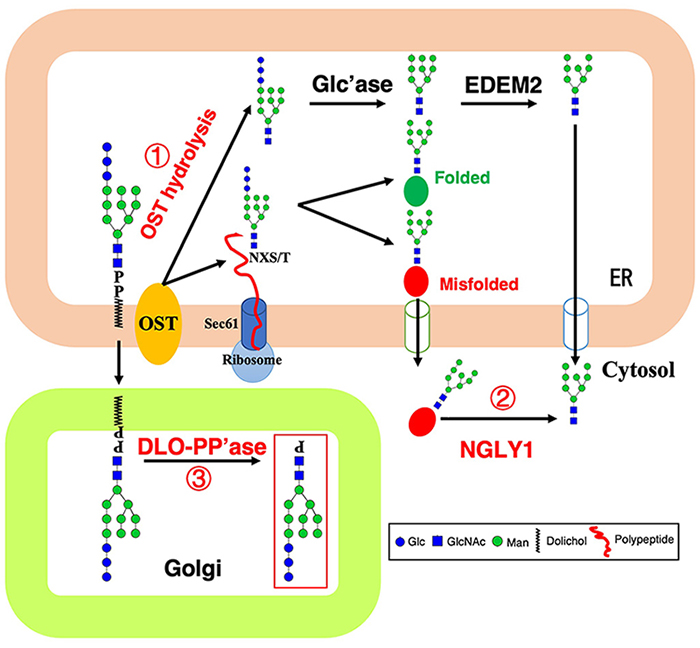
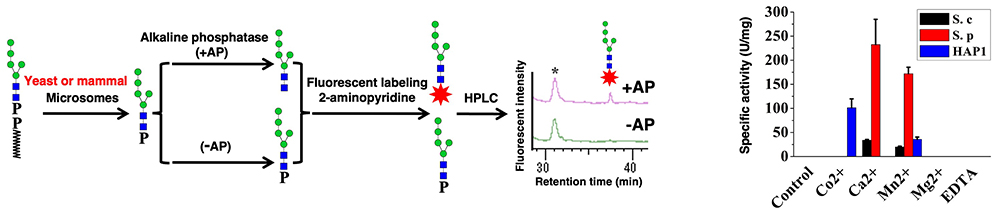
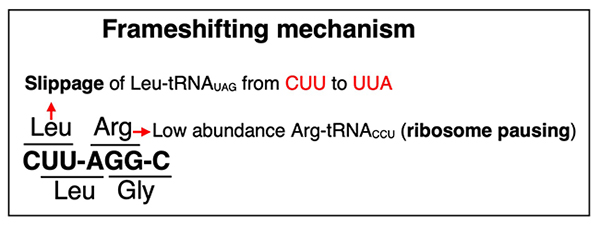
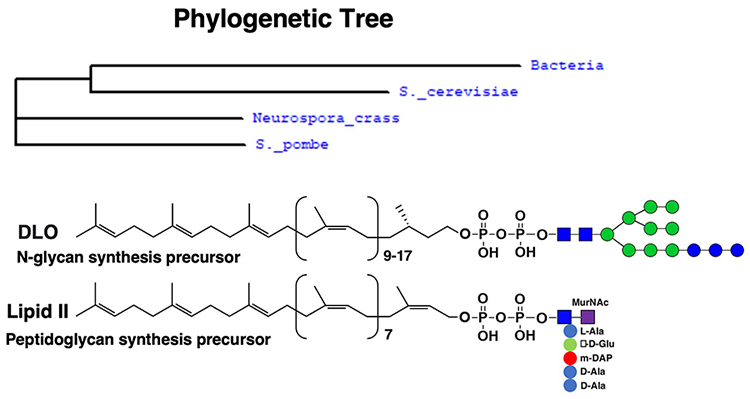
Aebi M. N-linked protein glycosylation in the ER. Biochimica et Biophysica Acta. 2013;1833(11):2430-7.
Wild R, Kowal J, Eyring J, Ngwa EM, Aebi M, Locher KP. Structure of the yeast oligosaccharyltransferase complex gives insight into eukaryotic N-glycosylation. Science. 2018;359(6375):545-50.
Bai L, Wang T, Zhao G, Kovach A, Li H. The atomic structure of a eukaryotic oligosaccharyltransferase complex. Nature. 2018;555(7696):328-33.
Hsu A-F, Baynes JW, Heath EC. The role of a dolichol-oligosaccharide as an intermediate in glycoprotein biosynthesis. Proceedings of the National Academy of Sciences of the United States of America. 1974;71(6):2391-5.
Harada Y, Hirayama H, Suzuki T. Generation and degradation of free asparagine-linked glycans. Cellular and Molecular Life Sciences. 2015;72(13):2509-33.
Cacan R, Hoflack B, Verbert A. Fate of oligosaccharide-lipid intermediates synthesized by resting rat-spleen lymphocytes. European Journal of Biochemistry. 1980;106(2):473-9.
Oliver GJ, Harrison J, Hemming FW. The mannosylation of dolichol-diphosphate oligosaccharides in relation to the formation of oligosaccharides and glycoproteins in pig-liver endoplasmic reticulum. European Journal of Biochemistry. 1975;58(1):223-9.
Hanover JA, Lennarz WJ. Transmembrane assembly of N-linked glycoproteins. Studies on the topology of saccharide synthesis. Journal of Biological Chemistry. 1982;257(6):2787-94.
Cacan R, Cecchelli R, Verbert A. Catabolic pathway of oligosaccharide-diphospho-dolichol: Study of the fate of the oligosaccharidic moiety in mouse splenocytes. European Journal of Biochemistry. 1987;166(2):469-74.
Cacan R, Lepers A, Belard M, Verbert A. Catabolic pathway of oligosaccharide-diphospho-dolichol: Subcellular sites of the degradation of the oligomannoside moiety. European Journal of Biochemistry. 1989;185(1):173-9.
Cacan R, Villers C, Bélard M, Kaiden A, Krag SS, Verbert A. Different fates of the oligosaccharide moieties of lipid intermediates. Glycobiology. 1992;2(2):127-36.
Hoflack B, Cacan R, Verbert A. Dolichol pathway in lymphocytes from rat spleen. Influence of the glucosylation on the cleavage of dolichyl diphosphate oligosaccharides into phosphooligosaccharides. European Journal of Biochemistry. 1981;117(2):285-90.
Belard M, Cacan R, Verbert A. Characterization of an oligosaccharide-pyrophosphodolichol pyrophosphatase activity in yeast. Biochemical Journal. 1988;255(1):235-42.
Massarweh A, Bosco M, Iatmanen-Harbi S, Tessier C, Auberger N, Busca P, et al. Demonstration of an oligosaccharide-diphosphodolichol diphosphatase activity whose subcellular localization is different than those of dolichyl-phosphate-dependent enzymes of the dolichol cycle. Journal of Lipid Research. 2016;57(6):1029-42.
Massarweh A, Bosco M, Iatmanen-Harbi S, Tessier C, Amana L, Busca P, et al. Brefeldin A promotes the appearance of oligosaccharyl phosphates derived from Glc3Man9GlcNAc2-PP-dolichol within the endomembrane system of HepG2 cells. Journal of Lipid Research. 2016;57(8):1477-91.
Peric D, Durrant-Arico C, Delenda C, Dupré T, De Lonlay P, de Baulny HO, et al. The compartmentalisation of phosphorylated free oligosaccharides in cells from a CDG Ig patient reveals a novel ER-to-cytosol translocation process. Plos One. 2010;5(7):e11675.
Vleugels W, Duvet S, Peanne R, Mir A-M, Cacan R, Michalski J-C, et al. Identification of phosphorylated oligosaccharides in cells of patients with a congenital disorders of glycosylation (CDG-I). Biochimie. 2011;93(5):823-33.
Harada Y, Nakajima K, Masahara-Negishi Y, Freeze HH, Angata T, Taniguchi N, et al. Metabolically programmed quality control system for dolichol-linked oligosaccharides. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(48):19366-71.
Harada Y, Nakajima K, Suzuki T, Fukushige T, Kondo K, Seino J, et al. Glycometabolic regulation of the biogenesis of small extracellular vesicles. Cell Reports. 2020;33(2):108261.
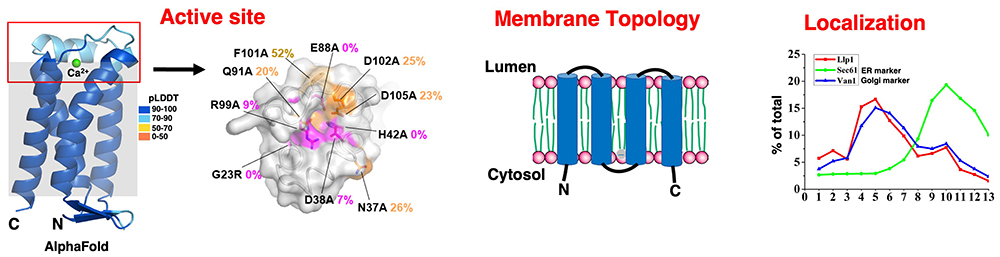
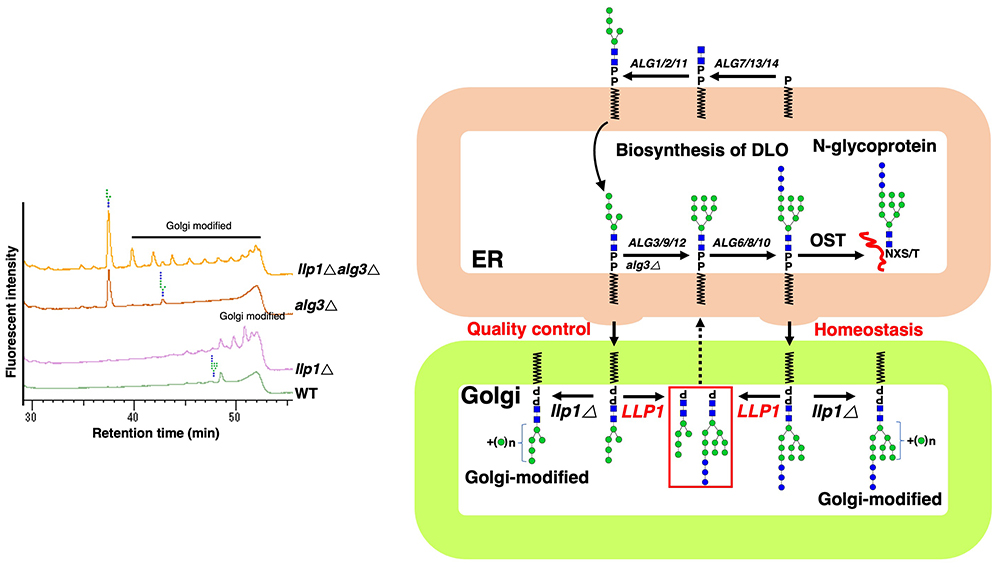
Li S-T, Kamada K, Honda A, Seino J, Matsuda T, Suzuki T, et al. LLP1 is a pyrophosphatase involved in homeostasis/quality control of dolichol-linked oligosaccharide. Journal of Cell Biology. 2025;224(11):e202501239.
Xu H, Boeke JD. Host genes that influence transposition in yeast: the abundance of a rare tRNA regulates Ty1 transposition frequency. Proceedings of the National Academy of Sciences of the United States of America. 1990;87(21):8360-4.
Kawakami K, Pande S, Faiola B, Moore DP, Boeke JD, Farabaugh PJ, et al. A rare tRNA-Arg (CCU) that regulates Ty1 element ribosomal frameshifting is essential for Ty1 retrotransposition in Saccharomyces cerevisiae. Genetics. 1993;135(2):309-20.
Belcourt MF, Farabaugh PJ. Ribosomal frameshifting in the yeast retrotransposon Ty: tRNAs induce slippage on a 7 nucleotide minimal site. Cell. 1990;62(2):339-52.
Sundararajan A, Michaud WA, Qian Q, Stahl G, Farabaugh PJ. Near-cognate peptidyl-tRNAs promote +1 programmed translational frameshifting in yeast. Molecular Cell. 1999;4(6):1005-15.
Arthur M, Molinas C, Depardieu F, Courvalin P. Characterization of Tn1546, a Tn3-related transposon conferring glycopeptide resistance by synthesis of depsipeptide peptidoglycan precursors in Enterococcus faecium BM4147. Journal of Bacteriology. 1993;175(1):117-27.
Arthur M, Depardieu F, Molinas C, Reynolds P, Courvalin P. The vanZ gene of Tn1546 from Enterococcus faecium BM4147 confers resistance to teicoplanin. Gene. 1995;154(1):87-92.
Pootoolal J, Neu J, Wright GD. Glycopeptide antibiotic resistance. Annual Review of Pharmacology and Toxicology. 2002;42(1):381-408.
Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A, et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature. 2024; 630(8016):493-500.
Rip JW, Chaudhary N, Carroll K. The submicrosomal distribution of dolichyl phosphate and dolichyl phosphate phosphatase in rat liver. Journal of Biological Chemistry. 1983;258(24):14926-30.
Rip JW, Chaudhary N, Carroll KK. Distribution and metabolism of dolichol and dolichyl phosphate in rat liver. Canadian Journal of Biochemistry and Cell Biology. 1983;61(9):1025-31.