氏名:太田 隼人
創価大学糖鎖生命システム融合研究所、特任助教
2023年 創価大学大学院工学研究科生命情報工学専攻博士課程 修了、博士(工学)を取得。2023年9月から2025年9月まで創価大学理工学部 助教。2025年10月より現職。

氏名:山本 一夫
お茶の水女子大学ヒューマンライフサイエンス研究所 客員教授
1979年 東京大学薬学部卒、1984年 東京大学大学院薬学系研究科博士課程修了(薬学博士)、1984年 東京都臨床医学総合研究所 研究員、1986年 東京大学薬学部助手、1997年 東京大学大学院薬学系研究科 助教授、1999年 東京大学大学院新領域創成科学研究科 教授、2022年 東京大学名誉教授、2022年 千葉大学大学院医学研究院 特任教授、2024年から現在に至る。

氏名:西原 祥子
創価大学糖鎖生命システム融合研究所、所長、特別教授
1982年 東京大学大学院理学系研究科化学専攻博士課程 修了、理学博士取得。慶応義塾大学医学部薬化学研究所、東京慈恵会医科大学、ノースキャロライナ大学、三菱化成生命科学研究所を経て、1991年より創価大学。現在、日本糖鎖科学コンソーシアム (JCGG)会長、Glycoconjugate Journal のEditorial Boardメンバーを務めている。
研究テーマ:多能性幹細胞における糖鎖機能の網羅的解析。ショウジョウバエを用いた糖鎖機能の解析。糖鎖が関わる未診断疾患や希少疾患の解析。
前立腺がんはアンドロゲン受容体(AR)シグナルに依存して増殖するが、ホルモン除去療法後に一部の症例が去勢抵抗性前立腺がん(CRPC)へと進展する。その分子基盤として、ARを介さない「バイパスシグナル」の活性化が知られているが、そのシグナル制御機構は十分に理解されていない。本稿では、細胞表面に存在する糖鎖、特に硫酸化グリコサミノグリカン(GAG)に着目し、3-O-硫酸化ヘパラン硫酸(3-OS HS)によるEGFRシグナルの活性化と、コンドロイチン硫酸E(CS-E)によるIL-6/STAT3シグナルの増強という二つの独立した去勢抵抗性獲得機構を概説し、新たな治療標的としてのGAGの可能性を示す。
前立腺がんはアンドロゲン受容体(androgen receptor:AR)シグナルに依存して増殖する男性における代表的ながんであり、初期治療としてアンドロゲン除去療法が広く用いられている。しかし、一部の症例において腫瘍は治療抵抗性を獲得し、去勢抵抗性前立腺がん(castration-resistant prostate cancer:CRPC)へと進展する1。CRPCではARシグナルの再活性化に加え、ARを介さない代替的増殖経路、いわゆる「バイパスシグナル」が重要な役割を果たすと考えられているが、その統合的な制御機構はいまだ十分に理解されていない2-4。
近年、がん研究および臨床診断の分野では、タンパク質量のみならず、それに付加された糖鎖構造が疾患状態や悪性度を反映することが明らかになりつつある。前立腺がんにおいても、診断マーカーとして用いられる前立腺特異抗原(prostate specific antigen:PSA)の糖鎖構造に着目し、診断精度を向上させる研究も進んでいる5。一方で、糖鎖は単なる分子マーカーにとどまらず、細胞表面において受容体とリガンドの相互作用を調節し、細胞内シグナル伝達を制御する機能分子として働くことが知られている6,7。特にヘパラン硫酸やコンドロイチン硫酸に代表される硫酸化グリコサミノグリカン(glycosaminoglycan:GAG)は、多様な増殖因子やサイトカインの結合を媒介することで、がん細胞の増殖、生存、環境応答に深く関与する8。
本稿では、前立腺がん細胞がホルモン除去環境に適応する過程で生じる糖鎖構造の再編成に着目し、3-O-硫酸化ヘパラン硫酸(3-O-sulfated heparan sulfate:3-OS HS)によるEGF受容体(epidermal growth factor receptor:EGFR)シグナル活性化と、コンドロイチン硫酸E(chondroitin sulfate E:CS-E)によるinterleukin (IL)-6/signal transducer and activator of transcription(STAT)3シグナルの制御という二つの糖鎖依存的去勢抵抗性獲得機構について概説する。これらの知見を通して、糖鎖がCRPCにおけるシグナル選択性を規定する分子基盤として機能する可能性を議論する。
細胞は周囲環境からの情報を、細胞膜上に存在する受容体を介して受け取り、増殖、分化、生存といった多様な細胞応答を制御している。増殖因子やサイトカインなどのリガンドが受容体に結合することで細胞内シグナル伝達が開始されるが、この相互作用は必ずしも受容体とリガンドのみで完結するものではない。細胞表面に存在する糖鎖が受容体複合体の形成やリガンド結合効率を調節し、「共受容体」として機能することがある6,7。
糖鎖の中でも、GAGは、細胞表面および細胞外マトリックスに豊富に存在する直鎖状多糖である。GAGは、N-アセチルグルコサミンあるいはN-アセチルガラクトサミンといったアミノ糖と、グルクロン酸やイズロン酸などのウロン酸からなる二糖単位の繰り返し構造を基本としており、ヘパラン硫酸(heparan sulfate:HS)、コンドロイチン硫酸(chondroitin sulfate:CS)、デルマタン硫酸(dermatan sulfate:DS)、ケラタン硫酸(keratan sulfate:KS)などに分類される。
GAGの機能的特徴は、糖鎖骨格そのものよりも硫酸基の付加位置や数、すなわち硫酸化パターンによって規定される点にある。硫酸化様式の違いはリガンドとの結合特異性を決定し、同一種類のGAGであっても活性化される受容体や下流シグナル経路が大きく異なる。このような硫酸化パターンは、細胞が周囲の多様な情報を選択的に受容するための分子基盤として機能する。したがってGAGは単なる細胞表面の構造成分ではなく、細胞外シグナルの受容効率や選択性を調節する能動的な制御分子として理解することができる。
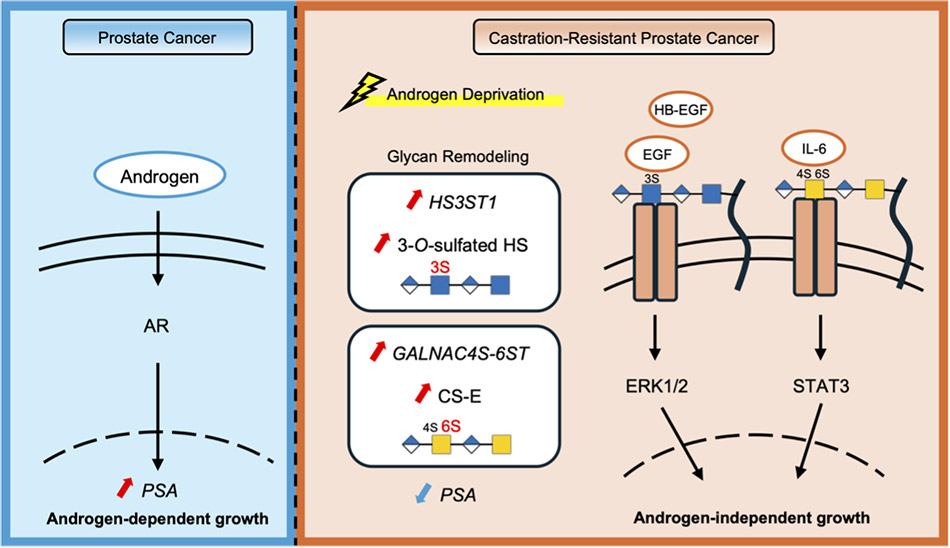
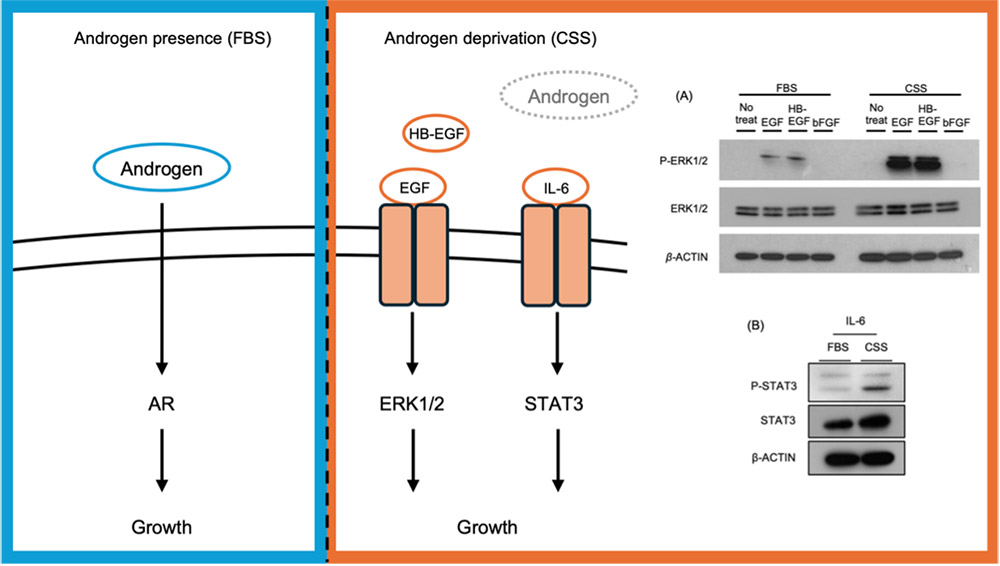
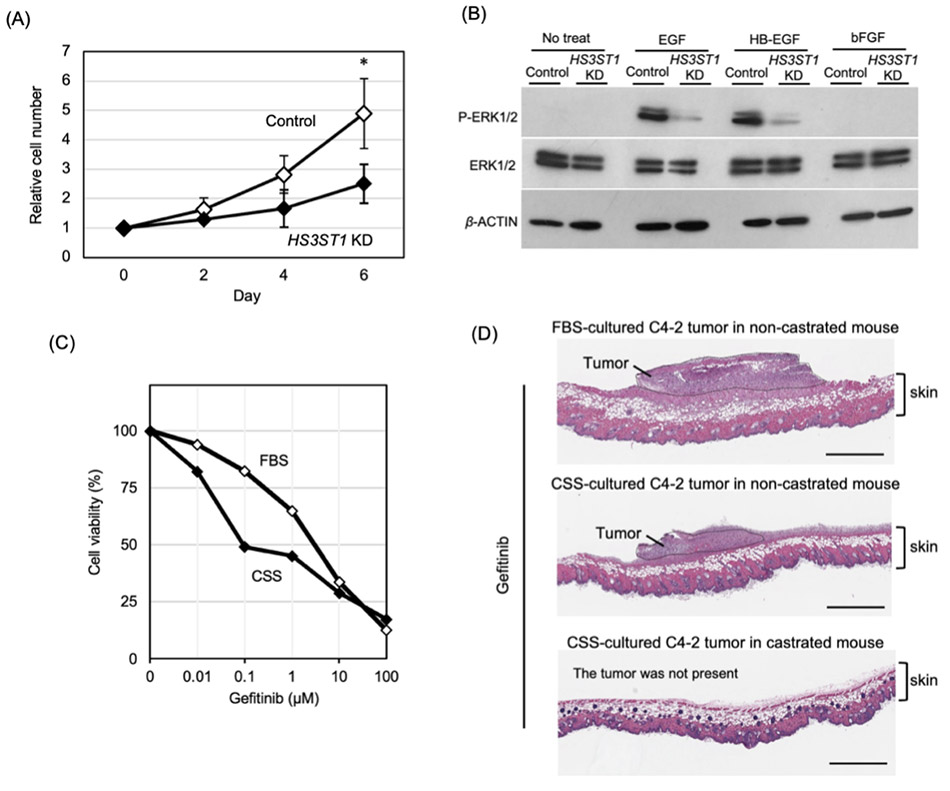
CRPC細胞株であるC4-2細胞はホルモン存在条件(Fetal bovine serum: FBS)とホルモン除去条件(Charcoal stripped serum: CSS)の両培養条件において増殖することができる9-11。FBSおよびCSS含有培地で培養した際のシグナルの変化を検討すると、CSS培養条件下ではepidermal growth factor(EGF)とheparin binding(HB)-EGFによって活性化するEGFRシグナルと、IL-6/STAT3シグナルがARシグナルの代わりに活性化することが示された(図 1)。これらのシグナルがアンドロゲン非依存的なバイパスシグナルとして機能することはこれまでの報告からも明らかであるが、その活性化メカニズムについての詳細は不明であった。
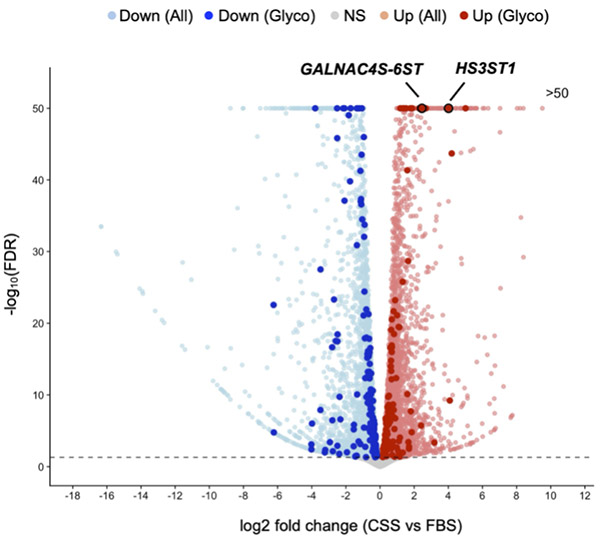
ホルモン除去という環境変化に応答して前立腺がんにおけるGAGの硫酸化状態が変化し、それがAR非依存的なCRPCの増殖の一因となっていることが明らかになってきた10,11。C4-2細胞をFBSとCSSで培養した際の遺伝子発現の比較(RNA-seq解析)により、多数の遺伝子の発現が変化することが明らかになった(図 2)。ホルモン除去によって、糖鎖関連遺伝子のうち131遺伝子が有意に上昇、151遺伝子が有意に減少し、ダイナミックに発現変動することが示された。特に、GAGを合成するヘパラン硫酸3-O-硫酸転移酵素 HS3ST1およびCS-E合成酵素GALNAC4S-6STの発現上昇が特徴的であった。これらの糖鎖関連遺伝子の発現変化がシグナル活性に影響を与えるならば、ホルモン除去が単に転写ネットワークを変化させるだけでなく、細胞表面糖鎖の発現を再構築することを示唆している。すなわちCRPC細胞では、糖鎖関連遺伝子の発現制御を通じて、AR非依存的シグナル伝達を活性化する細胞表面環境が形成されると考えられた。
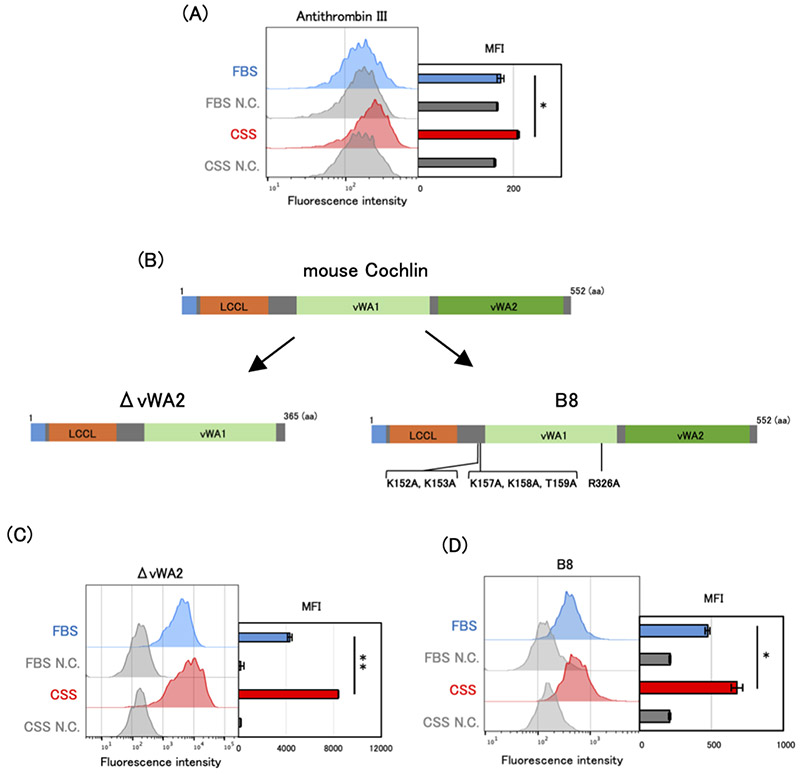
硫酸化GAGは、その構造多様性と解析の困難さから、機能解析が制限されてきた分子群である。特に、細胞表面に存在する特定の硫酸化パターンを選択的に識別することは容易ではなく、従来の抗体やレクチンでは十分な特異性が得られない場合が多かった。近年、硫酸化糖鎖に結合するレクチンであるCochlinを基盤として、特定の硫酸化構造を選択的に認識する人工改変体が開発されてきた11-13。これらの人工改変Cochlinは、高度に硫酸化された糖鎖構造に対して高い親和性を示し、フローサイトメトリーや組織染色などを通じて細胞表面に存在する硫酸化糖鎖を直接可視化する手段を提供する。
マウスCochlinは主にヘパリンおよびCS-Eを認識するレクチンであり、LCCLドメイン、vWA1ドメイン、vWA2ドメインの3つのドメインから構成されている。vWA2ドメインを欠失したΔvWA2 Cochlinは、野生型Cochlinと比較してより広範な硫酸化GAGを認識する特性を示す。一方、6つのアミノ酸置換(K152A、K153A、K157A、K158A、T159A、R236A)を導入したB8 Cochlinは、野生型と比較してヘパリンおよびCS-Eに対する特異性と親和性が向上した人工改変体である(図 3)。
本稿で紹介する3-OS HSおよびCS-Eに関する研究では、このようなCochlinを基盤とした糖鎖認識ツールが重要な役割を果たした10,11。具体的には、FBSおよびCSS条件で培養したC4-2細胞の細胞表面を人工改変Cochlinで染色し、フローサイトメトリーにより解析した。その結果、3-OS HSを含む硫酸化GAGを認識するΔvWA2 Cochlinでは、ホルモン除去条件(CSS)で染色強度が増加した(図 3)。さらに HS3ST1をノックダウンすると染色が低下し、また3-OS HSを特異的に認識するAntithrombin IIIによる染色でも同様の傾向が観察された(図 3A-C)。これらの結果から、ホルモン枯渇環境では3-OS HSを含む硫酸化GAGが増加することがわかった。同様に、CS-Eを高親和性で認識するB8 CochlinでもCSS条件で染色強度の増加が認められたことから、ホルモン除去環境下ではCS-Eの細胞表面発現が増加することが示された(図 3D)。このような糖鎖構造を認識する分子ツールの発展は、糖鎖研究を進展させる重要な原動力となっている。
先述したように、CRPCではヘパラン硫酸3-O-硫酸転移酵素 HS3ST1の発現上昇に伴い、3-OS HS構造が増加することが明らかとなった。HS3STファミリーによって合成される3-O-硫酸化は細胞内シグナル伝達を制御する14-16。そこで、HS3ST1による3-OS HS構造の増加がCRPCにおけるAR非依存的増殖に関与するかを検討するため、 HS3ST1遺伝子のノックダウンを行った10。 HS3ST1をノックダウンしたC4-2細胞では細胞増殖能が低下し、EGFRシグナルの活性が著しく抑制された(図 4A,B)。この結果は、CRPCにおいて3-OS HSがEGFRシグナルの活性化を介して細胞増殖を促進していることを示していた。
さらに、EGFR阻害剤Gefitinibを用いて増殖抑制効果を比較したところ、ホルモン存在下(FBS)条件で培養したC4-2細胞に比べ、ホルモン枯渇(CSS)条件で培養した細胞において、より強い増殖抑制効果が認められた(IC50:FBS 3.67 μM、CSS 0.33 μM)(図 4C)。この結果は、ホルモン除去環境下においてCRPC細胞がEGFRシグナルへの依存性を高めていることを示している。FBSまたはCSS条件で培養したC4-2細胞を通常マウスおよび去勢マウスの皮下に移植しGefitinibを投与したところ、よりアンドロゲン除去環境に近い条件で形成された腫瘍において、強い腫瘍増殖抑制効果が観察された(図 4D)。これらの結果から、ホルモン枯渇環境においてEGFR依存的な増殖機構がより重要になることが明らかになった。
本機構の分子基盤として、3-O-硫酸化を受けたHSがEGFやHB-EGFなどのEGFRリガンドの受容体結合を促進し、EGFRシグナルを効率的に活性化する可能性が考えられた。重要な点は、このシグナル活性化がリガンドや受容体の発現量の変化によってではなく、細胞表面糖鎖の硫酸化構造の変化によって引き起こされる点である。
これらの結果は、ホルモン除去環境下において3-OS HSがEGFRシグナルの活性化を促進し、CRPC細胞がAR依存的増殖からEGFR依存的増殖へとシグナル依存性を再構築することを示している。したがって、HSの3-O-硫酸化は、CRPCにおけるAR非依存的増殖を促進する分子基盤の一つであると考えられた。
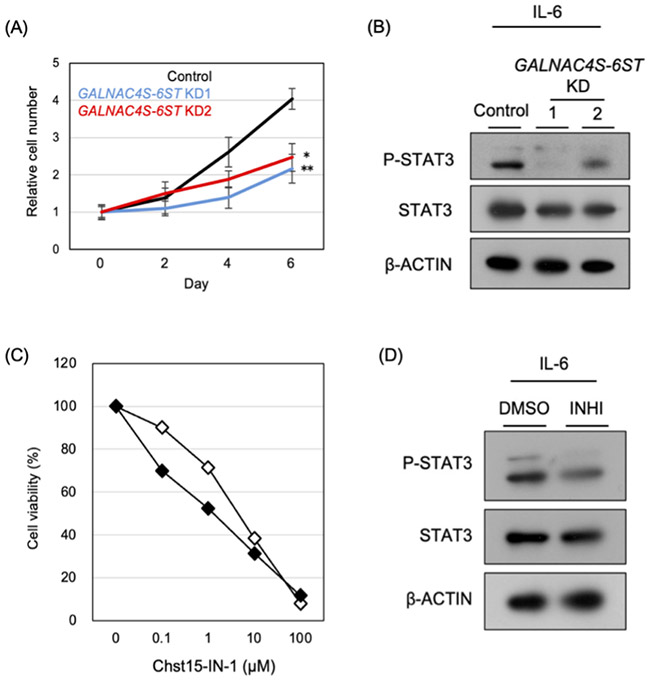
C4-2細胞をホルモン枯渇(CSS)条件で培養すると、CS-Eの合成酵素であるGALNAC4S-6STの発現上昇に伴い、CS-E構造の細胞表面での発現が増加する。CS-Eも種々の細胞において、細胞内シグナル伝達を制御することが知られている17-21。
去勢抵抗性獲得過程におけるCS-Eの機能を検討するため、GALNAC4S-6STの発現を抑制したところ、C4-2細胞の増殖能が低下した(図 5A)11。このとき、IL-6/STAT3シグナルの活性化が抑制されることが示された(図 5B)。一方で、EGFRやWntなど他のシグナル経路には大きな影響は認められないことから、CS-EはCRPCにおいてIL-6/STAT3シグナルを選択的に制御する糖鎖構造であると考えられた。CS-EはIL-6の細胞表面への結合を促進し、IL-6受容体複合体の形成を安定化させることでSTAT3のリン酸化を増強すると考えられた。さらに、CS-Eの前駆体構造であるCS-AがCRPCの進展に関与することも報告されている22,23。これらの知見は、CS-Aの増加がCS-Eの産生増加につながり、その結果としてCS-Eを介した増殖機構の活性化に寄与している可能性を示唆した。
さらに、GALNAC4S-6ST阻害剤(Chst15-IN-1)をFBSおよびCSS含有培地で前培養したC4-2細胞に作用させたところ、CSS条件で培養した細胞においてより強い増殖抑制効果が認められた(IC50:FBS 4.46 μM、CSS 1.39 μM)(図 5C)。また、ノックダウン実験の結果と一致して、この阻害剤はSTAT3のリン酸化レベルも低下させた(図 5D)。
これらの結果から、ホルモン除去環境ではC4-2細胞がCS-E/IL-6/STAT3シグナルへの依存性を高め、この経路を介して増殖を維持していることが示唆された。すなわち、CRPCではCS-E依存的なIL-6/STAT3シグナルの活性化が促進され、CRPCのAR非依存的な増殖を支える重要な分子機構として機能していると考えられた。
本稿で紹介した二つの経路、すなわち3-OS HSを介したEGFRシグナルと、CS-Eを介したIL-6/STAT3シグナルは、いずれもCRPCにおいて活性化されるAR非依存的なバイパスシグナルである。両者は異なる受容体および下流経路を利用するものの、細胞表面糖鎖の硫酸化構造の変化によってシグナル伝達効率が制御されるという共通の特徴を持つ。重要な点は、これらのシグナル活性化がリガンドや受容体の発現量の増加によって引き起こされるのではなく、糖鎖構造の変化によって誘導されることである。すなわち、ホルモン除去という環境変化に応答して前立腺がん細胞は糖鎖合成および硫酸化経路を再編成し、増殖因子やサイトカインを効率よく捕捉できる細胞表面環境を形成する。その結果、ARシグナルが抑制された状況下でも、代替的な増殖・生存シグナルを維持することが可能になる。3-OS HSとCS-Eという二つの糖鎖修飾は、それぞれ異なるシグナル経路を支えるものの、がん細胞が環境変化に適応するための共通の分子戦略の一部を構成していると考えられた。
さらに、これら二つの糖鎖依存的シグナルは相互に排他的ではなく、同一細胞内で並行して機能しうる点も重要である。糖鎖構造の多様化はCRPCに多層的なシグナル冗長性をもたらしている可能性がある。この冗長性が、CRPC治療を困難にしている要因の一つであろう。
以上の知見は、糖鎖がCRPCにおけるシグナル選択性を決定する能動的な制御因子であることを示していた。糖鎖構造の変化という観点からシグナルネットワークを統合的に理解することは、CRPCの分子病態をより正確に捉え、新たな治療戦略の開発につながる可能性がある。