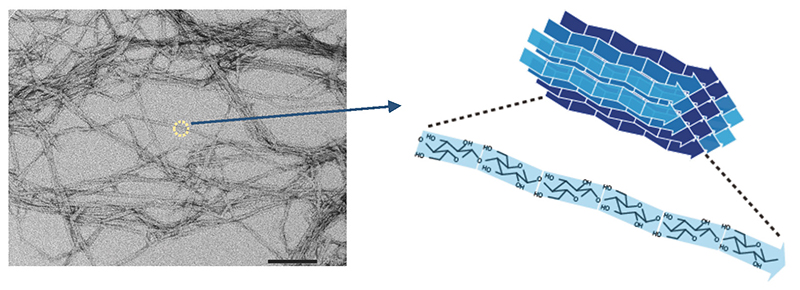
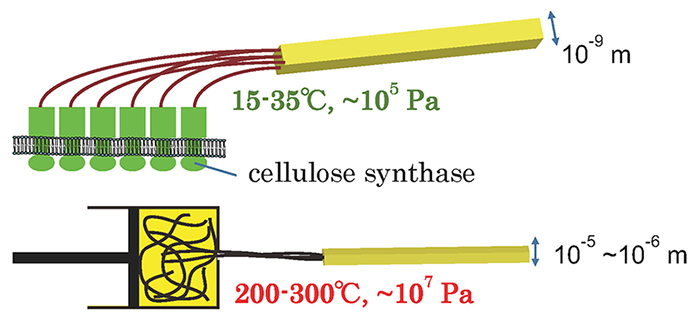
Sugiyama, J.; Horikawa, Y., The Cellulose Microfibril and Its Polymorphs. Mokuzai Gakkaishi2008, 54, (2), 49-57.
Wada, M.; Heux, L.; Sugiyama, J., Polymorphism of cellulose I family: Reinvestigation of cellulose IVl. Biomacromolecules2004, 5, (4), 1385-1391.
Atalla, R. H.; VanderHart, D. L., Native cellulose: A composite of two distinct crystalline forms. Science1984, 223, (4633), 283-285.
Sugiyama, J.; Vuong, R.; Chanzy, H., Electron Diffraction Study on the Two Crystalline Phases Occurring in Native Cellulose from an Algal Cell Wall. Macromolecules1991, 24, (14), 4168-4175.
Nishino, T.; Takano, K.; Nakamae, K., Elastic modulus of the crystalline regions of cellulose polymorphs. Journal of Polymer Science Part B: Polymer Physics1995, 33, (11), 1647-1651.
Kobayashi, S.; Hobson, L. J.; Sakamoto, J.; Kimura, S.; Sugiyama, J.; Imai, T.; Itoh, T., Formation and structure of artificial cellulose spherulites via enzymatic polymerization. Biomacromolecules2000, 1, (2), 168–173.
Fort, S.; Boyer, V.; Greffe, L.; Davies, G. J.; Moroz, O.; Christiansen, L.; Schülein, M.; Cottaz, S.; Driguez, H., Highly efficient synthesis of β(1 → 4)-oligo- and -polysaccharides using a mutant cellulase. Journal of the American Chemical Society2000, 122, (23), 5429-5437.
Hiraishi, M.; Igarashi, K.; Kimura, S.; Wada, M.; Kitaoka, M.; Samejima, M., Synthesis of highly ordered cellulose II in vitro using cellodextrin phosphorylase. Carbohydrate research2009, 344, (18), 2468-2473.
Yataka, Y.; Sawada, T.; Serizawa, T., Multidimensional Self-Assembled Structures of Alkylated Cellulose Oligomers Synthesized via in Vitro Enzymatic Reactions. Langmuir2016, 32, (39), 10120-10125.
Glaser, L., The enzymic synthesis of cellulose by Acetobacter xylinum. BBA - Biochimica et Biophysica Acta1957, 25, (2), 436.
Wong, H. C.; Fear, A. L.; Calhoon, R. D.; Eichinger, G. H.; Mayer, R.; Amikam, D.; Benziman, M.; Gelfand, D. H.; Meade, J. H.; Emerick, A. W.; Bruner, R.; Ben-Bassat, A.; Tal, R., Genetic organization of the cellulose synthase operon in Acetobacter xylinum. Proceedings of the National Academy of Sciences of the United States of America1990, 87, (20), 8130-8134. (in Japanese)
Saxena, I. M.; Lin, F. C.; Brown Jr, R. M., Cloning and sequencing of the cellulose synthase catalytic subunit gene of Acetobacter xylinum. Plant Molecular Biology1990, 15, (5), 673-683.
Imai, T., Challenging cellulose biosynthesis with a bacterial model. Cellulose Communications2011, 18, 156-162.
Bureau, T. E.; Brown Jr, R. M., In vitro synthesis of cellulose II from a cytoplasmic membrane fraction of Acetobacter xylinum. Proceedings of the National Academy of Sciences1987, 84, (20), 6985-9.
Morgan, J. L. W.; Strumillo, J.; Zimmer, J., Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature2013, 493, (7431), 181-186.
Hashimoto, A.; Shimono, K.; Horikawa, Y.; Ichikawa, T.; Wada, M.; Imai, T.; Sugiyama, J., Extraction of cellulose-synthesizing activity of Gluconacetobacter xylinus by alkylmaltoside. Carbohydrate Research2011, 346, (17), 2760–2768.
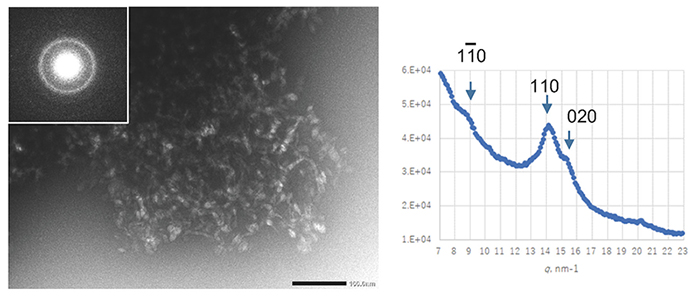
Penttilä, P. A.; Sugiyama, J.; Imai, T., Effects of reaction conditions on cellulose structures synthesized in vitro by bacterial cellulose synthases. Carbohydrate Polymers2016, 136, 656-666.
Ono, Y.; Tanaka, R.; Funahashi, R.; Takeuchi, M.; Saito, T.; Isogai, A., SEC–MALLS analysis of ethylenediamine-pretreated native celluloses in LiCl/N,N-dimethylacetamide: softwood kraft pulp and highly crystalline bacterial, tunicate, and algal celluloses. Cellulose2016, 23, (3), 1639-1647.
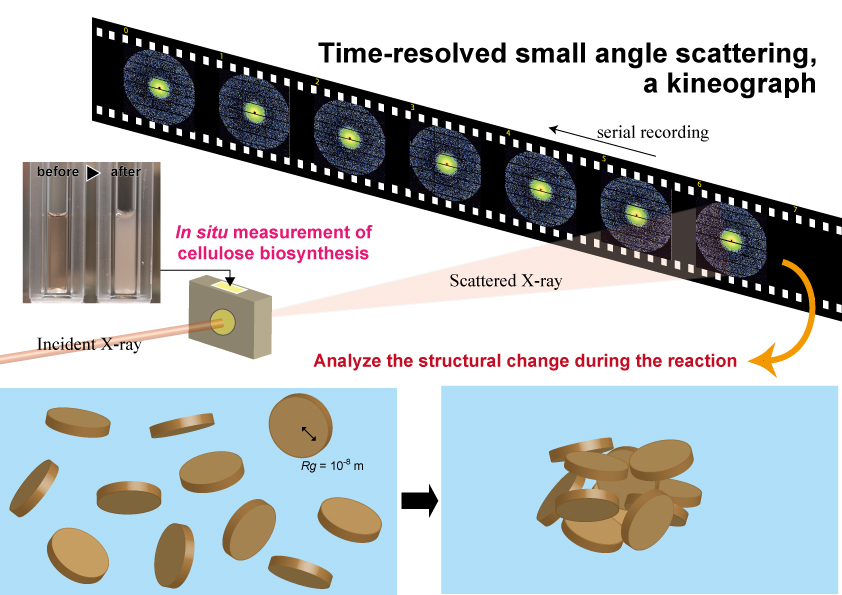
Tajima, H.; Penttilä, P.; Imai, T.; Yamamoto, K.; Yuguchi, Y., Observation of in vitro cellulose synthesis by bacterial cellulose synthase with time-resolved small angle X-ray scattering. International Journal of Biological Macromolecules2019, 130, (1), 765-777.
Omadjela, O.; Narahari, A.; Strumillo, J.; Melida, H.; Mazur, O.; Bulone, V.; Zimmer, J., BcsA and BcsB form the catalytically active core of bacterial cellulose synthase sufficient for in vitro cellulose synthesis. Proceedings of the National Academy of Sciences of the United States of America2013, 110, (44), 17856-17861.
Imai, T.; Sun, S. J.; Horikawa, Y.; Wada, M.; Sugiyama, J., Functional reconstitution of cellulose synthase in Escherichia coli. Biomacromolecules2014, 15, (11), 4206-4213.
Updegraff, D. M., Semimicro determination of cellulose inbiological materials. Analytical Biochemistry1969, 32, (3), 420-424.
Nakai, T.; Sugano, Y.; Shoda, M.; Sakakibara, H.; Oiwa, K.; Tuzi, S.; Imai, T.; Sugiyama, J.; Takeuchi, M.; Yamauchi, D.; Mineyukia, Y., Formation of highly twisted ribbons in a carboxymethylcellulase gene-disrupted strain of a cellulose-producing bacterium. Journal of Bacteriology2013, 195, (5), 958-964. (in Japanese)
Sunagawa, N.; Fujiwara, T.; Yoda, T.; Kawano, S.; Satoh, Y.; Yao, M.; Tajima, K.; Dairi, T., Cellulose complementing factor (Ccp) is a new member of the cellulose synthase complex (terminal complex) in Acetobacter xylinum. Journal of Bioscience and Bioengineering2013, 115, (6), 607-612.
Kimura, S.; Laosinchai, W.; Itoh, T.; Cui, X.; Linder, C. R.; Malcolm Brown Jr, R., Immunogold labeling of rosette terminal cellulose-synthesizing complexes in the vascular plant Vigna angularis. Plant Cell1999, 11, (11), 2075-2085.
Kimura, S.; Chen, H. P.; Saxena, I. M.; Brown R.M, Jr.; Itoh, T., Localization of c-di-GMP-binding protein with the linear terminal complexes of Acetobacter xylinum. Journal of Bacteriology2001, 183, (19), 5668-5674.
Kimura, S.; Nakashima, K., Recent progress in cellulose biosynthesis. Cellulose Communications2008, 15, (3), 109-113.
Sun, S. J.; Imai, T.; Sugiyama, J.; Kimura, S., CesA protein is included in the terminal complex of Acetobacter. Cellulose2017, 24, (5), 2017-2027.
Imai, T.; Sun, S.-j.; Horikawa, Y.; Wada, M.; Sugiyama, J., Functional reconstitution of cellulose synthase in Escherichia coli. Biomacromolecules2014, 15, (11), 4206–4213.
Nicolas, W. J.; Ghosal, D.; Tocheva, E. I.; Meyerowitz, E. M.; Jensen, G. J., Structure of the bacterial cellulose ribbon and its assembly-guiding cytoskeleton by electron cryotomography. Journal of Bacteriology2021, 203, (3), e00371-20.
Okano, T.; Sarko, A., Mercerization of cellulose. I. X‐ray diffraction evidence for intermediate structures. Journal of Applied Polymer Science 1984, 29, (12), 4175-4182.
Purushotham, P.; Ho, R.; Zimmer, J., Architecture of a catalytically active homotrimeric plant cellulose synthase complex. Science2020, 369, (6507), 1089-1094.