Atsushi Natsume
Currently, Dr. Natsume is Specially-appointed Professor, The Institute of Innovation for Future Society, Nagoya University. He graduated from Nagoya University School of Medicine in 1995 with an M.D. degree. His Ph.D. thesis research was carried out at Nagoya University and Aichi Cancer Center Research Institute in the cancer immune gene therapy research group of Professors Jun Yoshida and Toshitada Takahashi from 1997-1999. He did postdoctoral research in the Department of Molecular Genetics and Biochemistry at University of Pittsburgh, USA, from 1999-2002. Beginning in 2003, Dr. Natsume was a member of academic staff in the Department of Neurosurgery, Nagoya University and became an Associate Professor in 2007. Dr. Natsume has consistently been focused on multidisciplinary, problem-driven research aimed at elucidating the mechanism of malignant brain tumor evolution and creating next generation therapeutic and diagnostic approaches.
1. Introduction
We have focused on a molecule named podoplanin (PDPN) as a therapeutic target of malignant brain tumors. PDPN is a type I transmembrane mucin-like glycoprotein that is abundant in several solid tumors including squamous cell carcinoma, malignant mesothelioma, Kaposi sarcoma, angiosarcoma, testicular seminoma, and brain tumors. The expression level of PDPN is reported to increase as tumor malignancy increases. The side effects of currently available treatments can be avoided or mitigated if new measures are developed that only kill PDPN-expressing tumor cells.
2. The unique structure of PDPN
PDPN is a unique transmembrane glycoprotein receptor consisting of three domains: an amino terminal extracellular domain of approximately 130 amino acids, a single transmembrane domain of approximately 25 amino acids, and a short intracellular domain of approximately 10 amino acids (Figure 1). The extracellular domain is heavily glycosylated, a feature that has attracted the attention of many researchers in the field of glycoproteins. Lacking functional domains or enzymatic activities of its own, PDPN instead utilizes other proteins, including C-type lectin-like receptor-2 (CLEC-2), heat shock protein A9 (HSPA9), CD44, galectin 8, chemokine (C-C motif) ligand 21 (CCL21), ezrin, moesin, protein kinase A (PKA), and cyclin dependent kinase 5 (CDK5), to affect cell behavior such as tumor cell migration, invasion, and metastasis1-4.
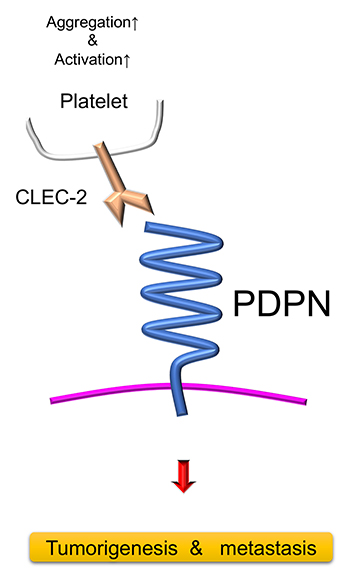
Figure 1. The structure and function of podoplanin (PDPN)
The long extracellular domain of PDPN is highly glycosylated and performs many molecular functions when interacted with other molecules. On the other hand, the intracellular domain is much shorter and non-functional. PDPN binds to platelet-derived C-type lectin-like receptor-2 (CLEC-2), which induces platelet coagulation and activation. PDPN directly promotes cancer malignancy, and loosens peritumoral attachment of fibroblasts and keratinocytes. In this manner, PDPN indirectly enhances tumor metastasis and invasion.
3. PDPN as a cancer biomarker
PDPN is expressed in many types of cancer including squamous cell carcinomas in the head, neck, esophagus, lung, and uterine cervix, testicular seminoma, and malignant mesothelioma1,3. In particular, oral cancer exemplifies the utility of PDPN expression as a cancer biomarker. PDPN expression increases cancer cell migration, which can lead to increased metastasis. Accordingly, its expression correlates with decreased 5-year survival rates of patients with these cancers. Moreover, PDPN expression increases the risk of malignant transformation from precancerous oral lesions (oral leukoplakias) by 3 fold5
As well as in cancer cells, PDPN expression can be found in cancer associated fibroblasts (CAFs)1,3,5 and peritumoral basal keratinocytes6 that might contribute to cancer cell motility, invasion, and metastasis. The mechanism of action remains unelucidated, but platelet CLEC-2 interacts with the adhesion molecule, E-cadherin, causing CAF and keratinocytes to stick to the basal cell layer, while the CLEC-2 ligand, PDPN, if abundant, inhibits the CLEC-2 and E-cadherin pathway7
4. The PDPN extracellular domain as a therapeutic target
Preclinical studies indicate that the extracellular domain of PDPN can be a target of small molecular chemicals, lectins, antibodies, and chimeric antigen receptor T cells (CAR-T cells) and can thereby be used to combat cancer.
PDPN binds with CLEC-2 on platelets in the bloodstream to facilitate tumor embolism and hematogenous metastasis8,9 (Figure 1). A derivative of 4-O-benzoyl-3-methoxy-beta-nitrostyrene (BMNS), compound “2CP,” effectively suppresses PDPN-mediated platelet aggregation and tumor cell-induced platelet activation8. In addition to synthetic molecules, lectins may be used to target PDPN on transformed cells. For example, Maackia amurensis seed lectin (MASL) binds to PDPN on melanoma and oral squamous cell carcinoma cells to inhibit their motility and growth in vitro and in syngeneic and xenograft mouse models 10. These findings have led to a clinical trial of MASL as therapy against oral cancer in the US(NCT04188665, https://clinicaltrials.gov/ct2/show/NCT04188665).
5. NZ-1-based CAR-T cell therapy to glioblastoma
Glioblastoma (GBM) is the most common and lethal primary malignant brain tumor in adults and GBM patients have a 5-year overall survival rate of less than 10%. Chimeric antigen receptor (CAR) transduced T cells can recognize predefined tumor surface antigens independent of major histocompatibility complex (MHC) restriction (Figure 2). Several CARs have been generated against antigens expressed in GBM, including epidermal growth factor receptor variant III (EGFRvIII)11-13, human epidermal growth factor receptor 2 (HER2)14, and interleukin-13 receptor alpha 2 (IL13Rα215). In brain tumors, PDPN is overexpressed in accordance with tumor malignancy16. PDPN has been recognized as a potential therapeutic target of CAR-T cells to treat brain tumors, and we generated a third-generation CAR that targeted PDPN using NZ-1-based single chain variable fragments (scFv) of an anti-pan-PDPN antibody17. CAR-transduced peripheral blood monocytes (PBMCs) were immunologically evaluated by calcein-mediated cytotoxic assay, ELISA, tumor size, and overall survival (Figure 3). The generated CAR-T cells were specific and effective against PDPN-positive GBM cells in vitro. A calcein-based nonradioisotope cytotoxic assay indicated that PDPN-positive LN319 cells and U87MG glioma cells were lysed by these NZ-1-CAR-T cells in an effector/target (E/T) ratio-dependent manner. In contrast, specific lysis was not observed against PDPN-knockout (KO)-glioma cells. In addition, NZ-1-CAR-T cells co-cultured with PDPN expressing glioma cells released significantly more IFNγ than did mock-transduced T cells.
Systemic injection of the CAR-T cells into an immunodeficient mouse model inhibited the growth of intracranial glioma xenografts in vivo. GBM cells were implanted into an immunodeficient mouse brain. Seven days after tumor implantation, NZ-1-CAR-T cells or mock-transduced T cells were infused intravenously via the tail vein. The non-treated mice were infused with PBS alone, and intracranial tumor growth was evaluated by 3T-MRI. In approximately 60% of the mice treated with NZ-1-CAR-T cells, tumors grew markedly slower and the mice survived significantly longer than control groups. Thus, CAR-T cell therapy that targets PDPN shows promise as an adoptive immunotherapy for mesenchymal GBM18.
However, PDPN is also present in normal tissues19,20, such as the lymphatic endothelium and lung type I alveolar cells. This raises safety concerns regarding PDPN-targeted therapies and necessitates the development of a better approach to specifically target cancer cells to overcome the on-target-off-tumor effect.
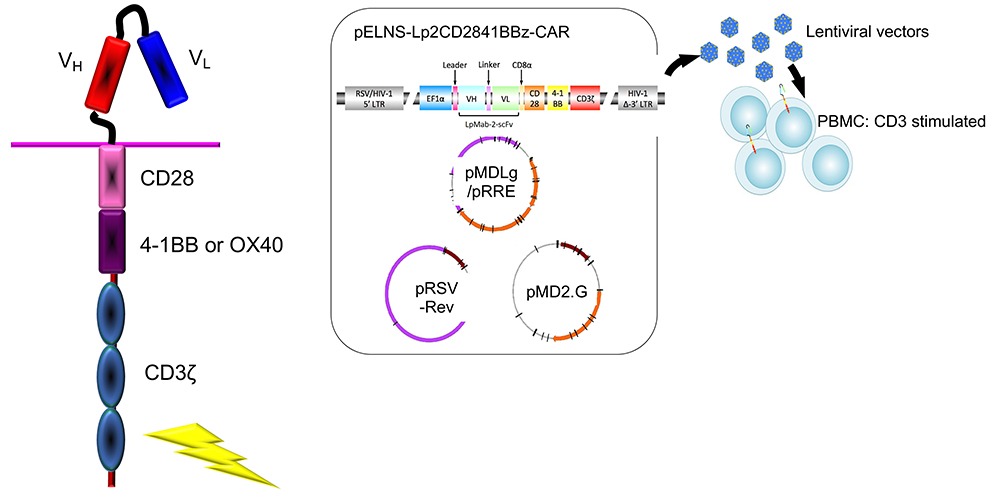
Figure 2. Generation of chimeric antigen receptor (CAR)
CARs consist of an extracellular domain derived from a single-chain variable fragment (scFv) taken from a tumor antigen-specific monoclonal antibody (mAb), a trans-membrane domain, and a cytoplasmic signaling domain CD3ζ chain (CD3ζ) derived from the T-cell receptor complex. CAR-transduced peripheral blood monocytes (PBMC) can recognize predefined tumor surface antigens independent of major histocompatibility complex (MHC) restriction. Third-generation CARs that include 2 costimulatory domains, such as CD28 and 4-1BB (CD137), have been described and are highly likely to lyse tumor cells.
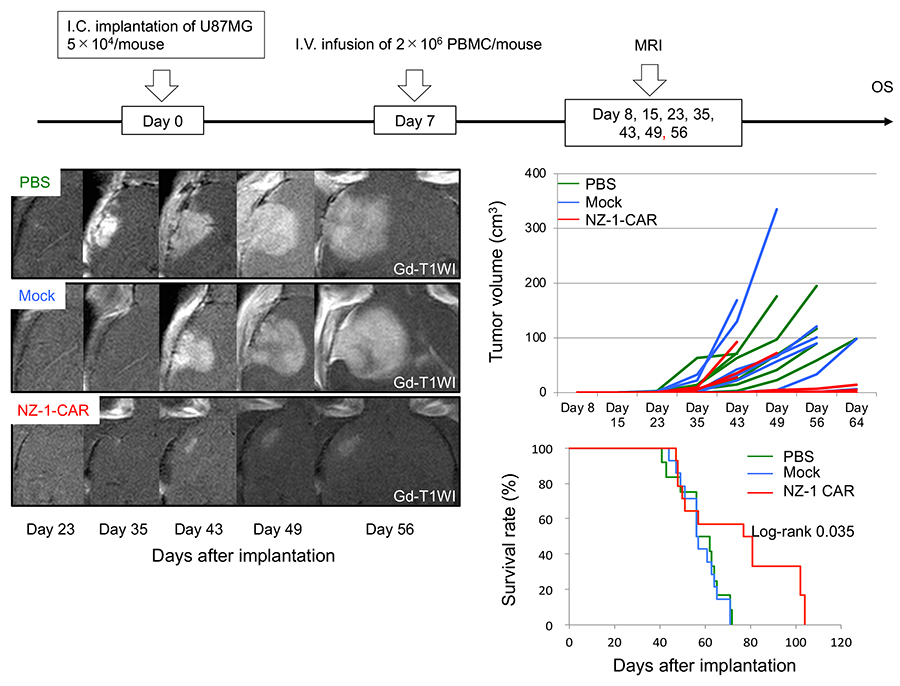
Figure 3. NZ-1-based chimeric antigen receptor T cells (CAR-T cells) suppress the growth of glioblastoma (GBM)
We constructed a lentiviral vector expressing a third-generation CAR comprised of a podoplanin (PDPN)-specific antibody (NZ-1-based single chain variable fragment [scFv]) with CD28, 4-1BB, and CD3ζ intracellular domains. CAR-transduced peripheral blood monocytes (PBMCs) were immunologically evaluated by calcein-mediated cytotoxic assay, ELISA, tumor size, and overall survival (OS). GBM cells were implanted into the brains of immunodeficient mice. Seven days after tumor implantation, NZ-1-CAR-T cells or mock-transduced T cells were infused intravenously (I.V.) via the tail vein (Upper). The non-treated mice were infused with PBS alone, and intracranial (I.C.) tumor growth was evaluated by 3T-MRI (Left). In approximately 60% of the mice treated with NZ-1-CAR-T cells, tumors grew markedly slower (Middle right) and the mice survived significantly longer than control groups (Lower right). Taken together, these data indicate that functionally active NZ-1-CAR-T cells recognize PDPN, enabling them to inhibit glioma cell growth and tumor progression.
6. Cancer-specific monoclonal antibody-based CAR-T cells to GBM
One concern with PDPN CAR-T cell therapy arises from nonspecific lysis of normal cells that express PDPN, including lymphatic endothelium and type I lung alveolar cells. Cancer-specific monoclonal antibodies (CasMabs) have been generated to address this concern.
One of our collaborative teams has generated a CasMab that recognizes aberrantly glycosylated PDPN in cancer cells, namely LpMab221,22. As Lp2 does not recognize PDPN-expressing normal cells, CAR-T cell therapy based on Lp2 can also be expected to be cancer-specific23. Lp2-CAR transduced T cells (Lp2-CAR-T) specifically targeted PDPN-expressing glioma cells, while sparing the PDPN-expressing normal cells (Figure 4).
7. A combination of CAR-T cell therapy with oncolytic herpes simplex virus
Next, we focused on combining CAR-T cell therapy with the very common and contagious herpes simplex virus (HSV). Symptoms of HSV infection include blisters on the lips or eyes. We have turned this virulent virus into an anti-cancer agent, a third-generation oncolytic recombinant HSV-1, designated G47Δ24. Its efficacy and safety was proved in a phase II clinical trial of this agent for GBM in Japan and G47Δ was approved by Ministry of Health, Labour and Welfare Japan as the first oncolytic therapy drug for malignant glioma in the world in June 202125. G47Δ not only killed solid tumors directly but also elicited an antitumor immune response. Combination therapy with Lp2-CAR-T cells and oncolytic virus G47Δ further inhibited tumor growth and improved survival. These findings suggest that the combination therapy of Lp2-CAR-T cells and G47Δ may be a promising approach to treat GBM23 (Figure 4).
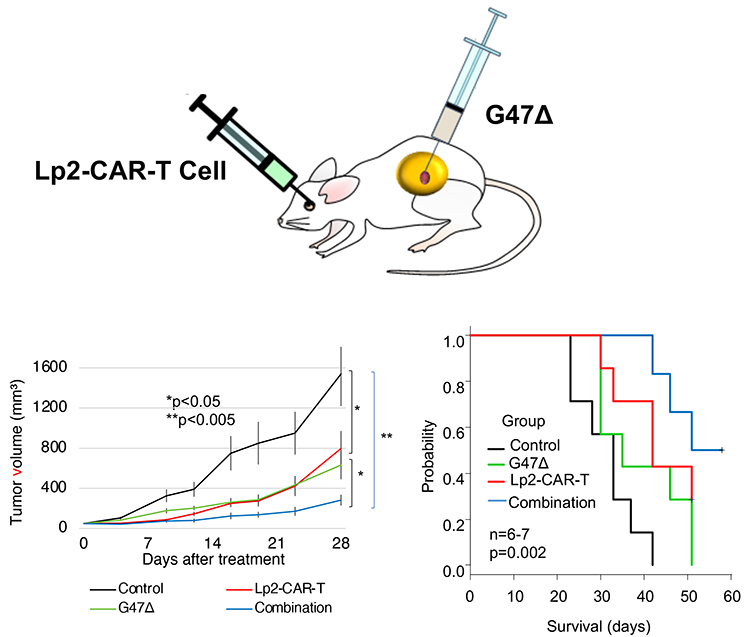
Figure 4. Efficacy of cancer-specific anti-podoplanin (PDPN) chimeric antigen receptor T cells (CAR-T cells) and oncolytic herpes virus G47Δ combination therapy against glioblastoma (GBM)
PDPN is expressed in various normal cells such as lymphatic endothelial cells, pulmonary alveolar type I cells, and podocytes. To overcome possible on-target-off-tumor effects, we produced a cancer-specific monoclonal antibody (CasMab, LpMab-2)-based chimeric antigen receptor (CAR). LpMab-2 (Lp2) reacted with PDPN-expressing cancer cells, but not normal cells. Systemic injection of Lp2-CAR-T cells inhibited the growth of a subcutaneous glioma xenograft model in immunodeficient mice. Combination therapy with Lp2-CAR-T and oncolytic virus G47Δ, a third-generation recombinant herpes simplex virus (HSV)-1, further inhibited the tumor growth and improved survival.
8. Conclusions
Cas-Mab-based CAR-T cell therapies may improve the safety and specificity of tumor-targeted therapies, while the expression of cancer-specific antigen in the tumor and normal tissues should be elaborately tested in the next preclinical trials. No adverse effects were observed in mice experimentally. However, it should be noted that this CAR is specific to human PDPN, and animal experiments cannot completely simulate its effect on human tissue. Moreover, it should also be noted that this is a limitation of animal experiments in general, and thorough safety precautions should be taken before any clinical application. Although a number of issues, such as the lack of complete response in most of the mouse models and the clarification of the exact immunological mechanisms involved in the response to the combination therapy, still need to be addressed, we have successfully established novel cancer-specific CAR-T cells against a promising tumor target antigen and have reported that it can be used in combination with an oncolytic virus against GBM.
References
Renart, J.; Carrasco-Ramírez, P.; Fernández-Muñoz, B.; Martín-Villar, E.; Montero, L.; Yurrita, M.M.; Quintanilla, M. New insights into the role of podoplanin in epithelial-mesenchymal transition. Int Rev Cell Mol Biol2015, 317, 185-239, doi:10.1016/bs.ircmb.2015.01.009.
Astarita, J.L.; Acton, S.E.; Turley, S.J. Podoplanin: emerging functions in development, the immune system, and cancer. Front Immunol2012, 3, 283, doi:10.3389/fimmu.2012.00283.
Wicki, A.; Christofori, G. The potential role of podoplanin in tumour invasion. Br J Cancer2007, 96, 1-5, doi:10.1038/sj.bjc.6603518.
Krishnan, H.; Retzbach, E.P.; Ramirez, M.I.; Liu, T.; Li, H.; Miller, W.T.; Goldberg, G.S. PKA and CDK5 can phosphorylate specific serines on the intracellular domain of podoplanin (PDPN) to inhibit cell motility. Exp Cell Res2015, 335, 115-122, doi:10.1016/j.yexcr.2015.04.019.
Cho, Z.; Konishi, E.; Kanemaru, M.; Isohisa, T.; Arita, T.; Kawai, M.; Tsutsumi, M.; Mizutani, H.; Takenaka, H.; Ozawa, T.; et al. Podoplanin expression in peritumoral keratinocytes predicts aggressive behavior in extramammary Paget's disease. J Dermatol Sci2017, 87, 29-35, doi:10.1016/j.jdermsci.2017.03.012.
Asai, J.; Hirakawa, S.; Sakabe, J.; Kishida, T.; Wada, M.; Nakamura, N.; Takenaka, H.; Mazda, O.; Urano, T.; Suzuki-Inoue, K.; et al. Platelets regulate the migration of keratinocytes via podoplanin/CLEC-2 signaling during cutaneous wound healing in mice. Am J Pathol2016, 186, 101-108, doi:10.1016/j.ajpath.2015.09.007.
Chang, Y.W.; Hsieh, P.W.; Chang, Y.T.; Lu, M.H.; Huang, T.F.; Chong, K.Y.; Liao, H.R.; Cheng, J.C.; Tseng, C.P. Identification of a novel platelet antagonist that binds to CLEC-2 and suppresses podoplanin-induced platelet aggregation and cancer metastasis. Oncotarget2015, 6, 42733-42748, doi:10.18632/oncotarget.5811.
Takemoto, A.; Miyata, K.; Fujita, N. Platelet-activating factor podoplanin: from discovery to drug development. Cancer Metastasis Rev2017, 36, 225-234, doi:10.1007/s10555-017-9672-2.
Hamilton, K.L.; Sheehan, S.A.; Retzbach, E.P.; Timmerman, C.A.; Gianneschi, G.B.; Tempera, P.J.; Balachandran, P.; Goldberg, G.S. Effects of Maackia amurensis seed lectin (MASL) on oral squamous cell carcinoma (OSCC) gene expression and transcriptional signaling pathways. J Cancer Res Clin Oncol2021, 147, 445-457, doi:10.1007/s00432-020-03456-8.
Kuramitsu, S.; Ohno, M.; Ohka, F.; Shiina, S.; Yamamichi, A.; Kato, A.; Tanahashi, K.; Motomura, K.; Kondo, G.; Kurimoto, M.; et al. Lenalidomide enhances the function of chimeric antigen receptor T cells against the epidermal growth factor receptor variant III by enhancing immune synapses. Cancer Gene Ther2015, 22, 487-495, doi:10.1038/cgt.2015.47.
Wang, S.; O'Rourke, D.M.; Chawla, S.; Verma, G.; Nasrallah, M.P.; Morrissette, J.J.D.; Plesa, G.; June, C.H.; Brem, S.; Maloney, E.; et al. Multiparametric magnetic resonance imaging in the assessment of anti-EGFRvIII chimeric antigen receptor T cell therapy in patients with recurrent glioblastoma. Br J Cancer2019, 120, 54-56, doi:10.1038/s41416-018-0342-0.
O'Rourke, D.M.; Nasrallah, M.P.; Desai, A.; Melenhorst, J.J.; Mansfield, K.; Morrissette, J.J.D.; Martinez-Lage, M.; Brem, S.; Maloney, E.; Shen, A.; et al. A single dose of peripherally infused EGFRvIII-directed CAR T cells mediates antigen loss and induces adaptive resistance in patients with recurrent glioblastoma. Sci Transl Med2017, 9, eaaa0984, doi:10.1126/scitranslmed.aaa0984.
Vitanza, N.A.; Johnson, A.J.; Wilson, A.L.; Brown, C.; Yokoyama, J.K.; Künkele, A.; Chang, C.A.; Rawlings-Rhea, S.; Huang, W.; Seidel, K.; et al. Locoregional infusion of HER2-specific CAR T cells in children and young adults with recurrent or refractory CNS tumors: an interim analysis. Nat Med2021, 27, 1544-1552, doi:10.1038/s41591-021-01404-8.
Brown, C.E.; Rodriguez, A.; Palmer, J.; Ostberg, J.R.; Naranjo, A.; Wagner, J.R.; Aguilar, B.; Starr, R.; Weng, L.; Synold, T.W.; et al. Off-the-shelf, steroid-resistant, IL13Rα2-specific CAR T cells for treatment of glioblastoma. Neuro Oncol2022, 24, 1318-1330, doi:10.1093/neuonc/noac024.
Eisemann, T.; Costa, B.; Harter, P.N.; Wick, W.; Mittelbronn, M.; Angel, P.; Peterziel, H. Podoplanin expression is a prognostic biomarker but may be dispensable for the malignancy of glioblastoma. Neuro Oncol2019, 21, 326-336, doi:10.1093/neuonc/noy184.
Chandramohan, V.; Bao, X.; Kato Kaneko, M.; Kato, Y.; Keir, S.T.; Szafranski, S.E.; Kuan, C.T.; Pastan, I.H.; Bigner, D.D. Recombinant anti-podoplanin (NZ-1) immunotoxin for the treatment of malignant brain tumors. Int J Cancer2013, 132, 2339-2348, doi:10.1002/ijc.27919.
Shiina, S.; Ohno, M.; Ohka, F.; Kuramitsu, S.; Yamamichi, A.; Kato, A.; Motomura, K.; Tanahashi, K.; Yamamoto, T.; Watanabe, R.; et al. CAR T cells targeting podoplanin reduce orthotopic glioblastomas in mouse brains. Cancer Immunol Res2016, 4, 259-268, doi:10.1158/2326-6066.CIR-15-0060.
Schacht, V.; Dadras, S.S.; Johnson, L.A.; Jackson, D.G.; Hong, Y.K.; Detmar, M. Up-regulation of the lymphatic marker podoplanin, a mucin-type transmembrane glycoprotein, in human squamous cell carcinomas and germ cell tumors. Am J Pathol 2005, 166, 913-921, doi:10.1016/S0002-9440(10)62311-5.
Kahn, H.J.; Marks, A. A new monoclonal antibody, D2-40, for detection of lymphatic invasion in primary tumors. Lab Invest2002, 82, 1255-1257, doi:10.1097/01.lab.0000028824.03032.ab.
Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci Rep2014, 4, 5924, doi:10.1038/srep05924.
Kaneko, M.K.; Yamada, S.; Nakamura, T.; Abe, S.; Nishioka, Y.; Kunita, A.; Fukayama, M.; Fujii, Y.; Ogasawara, S.; Kato, Y. Antitumor activity of chLpMab-2, a human-mouse chimeric cancer-specific antihuman podoplanin antibody, via antibody-dependent cellular cytotoxicity. Cancer Med2017, 6, 768-777, doi:10.1002/cam4.1049.
Chalise, L.; Kato, A.; Ohno, M.; Maeda, S.; Yamamichi, A.; Kuramitsu, S.; Shiina, S.; Takahashi, H.; Ozone, S.; Yamaguchi, J.; et al. Efficacy of cancer-specific anti-podoplanin CAR-T cells and oncolytic herpes virus G47∆ combination therapy against glioblastoma. Mol Ther Oncolytics2022, 26, 265-274, doi:10.1016/j.omto.2022.07.006.
Todo, T.; Ino, Y.; Ohtsu, H.; Shibahara, J.; Tanaka, M. A phase I/II study of triple-mutated oncolytic herpes virus G47∆ in patients with progressive glioblastoma. Nat Commun2022, 13, 4119, doi:10.1038/s41467-022-31262-y.
Frampton, J.E. Teserpaturev/G47∆: First Approval. BioDrugs2022, 36, 667-672, doi:10.1007/s40259-022-00553-7.