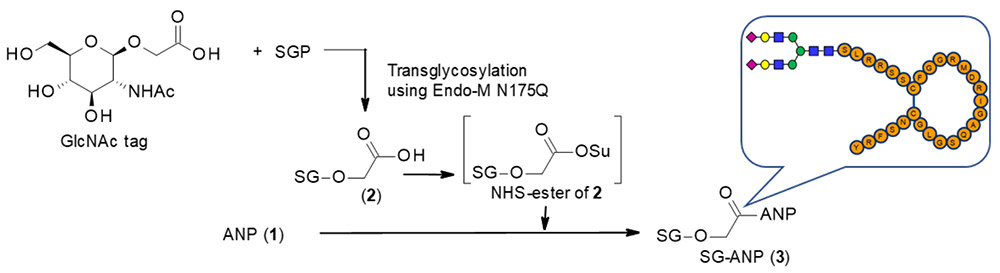
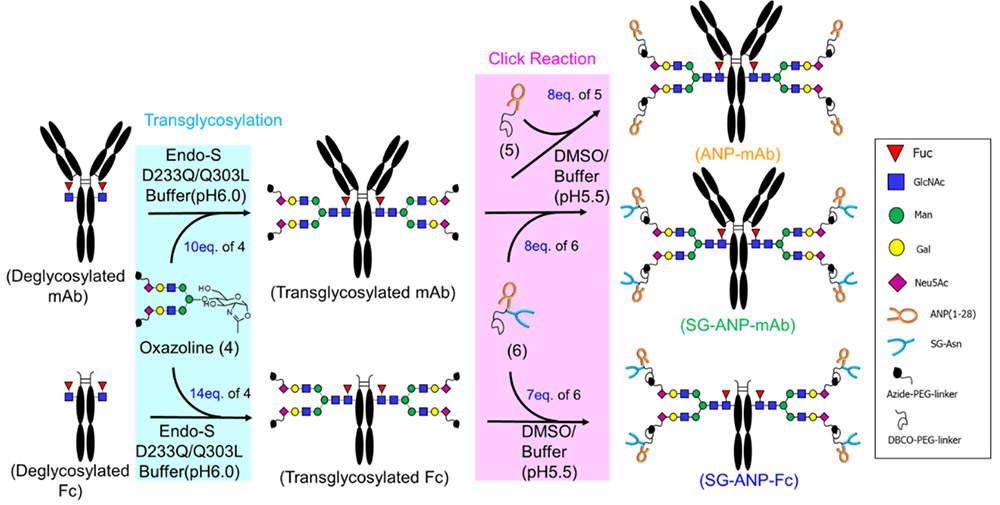
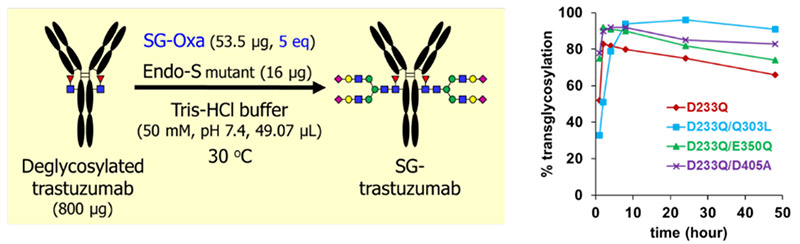
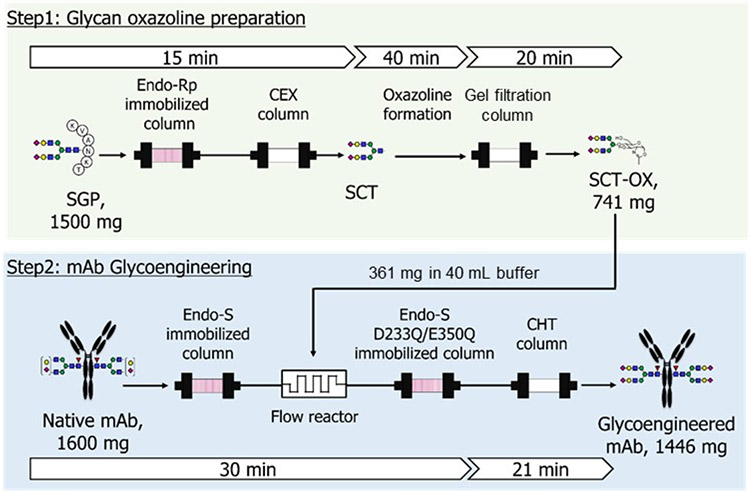
SGPにEndo M N175Qを作用させてGlcNAc tagへ糖鎖を転移させた後9,10、NHSエステル化し、ANP(1)と反応させてSG-ANP(3)を得た(図 1)。調製したSG-ANP(3)はラットにて強い薬理効果(グアニル酸シクラーゼA活性化)とネプリライシン(NEP)に対する代謝耐性を示した。
Li C, Wang LX. Chemoenzymatic methods for the synthesis of glycoproteins. Chem Rev. 2018; 118(17):8359-8413.
Nishiuchi Y, Elouali S, Noguchi M, Ochiai H. Conjugation of human N-glycans improves the drug properties of existing peptides and proteins. Chembiochem. 2025;26(8):e202401066.
Gimeno A, Ehlers AM, Delgado S, Langenbach JH, van den Bos LJ, Kruijtzer JAW, Guigas BGA, Boons GJ. Site-specific glyco-tagging of native proteins for the development of biologicals. J Am Chem Soc. 2024;146(50):34452-34465.
Trastoy B, Du JJ, García-Alija M, Li C, Klontz EH, Wang LX, Sundberg EJ, Guerin ME. Sculpting therapeutic monoclonal antibody N-glycans using endoglycosidases. Curr Opin Struct Biol. 2022;72:248-259.
Manabe S. Antibody glycoengineering for enhanced therapeutics: asymmetric glycan-homogeneous IgG and structurally homogeneous antibody-drug conjugate. Glycoforum. 2025;28(4):A16.
Matsuda Y, Chang JR, Mendelsohn BA. Advanced antibody-drug conjugates design: innovation in linker chemistry and site-specific conjugation technologies. Chembiochem. 2025;26(22):e202500305.
Iwamoto M, Yamaguchi T, Sekiguchi Y, Oishi S, Shiiki T, Soma M, Nakamura K, Yoshida M, Chaya H, Mori Y, Miyauchi R, Hasegawa J, Nagayama T, Honda T. Pharmacokinetic and pharmacodynamic profiles of glyco-modified atrial natriuretic peptide derivatives synthesized using chemo-enzymatic synthesis approaches. Bioconjug Chem. 2018;29(8):2829-2837.
Tomabechi Y, Suzuki R, Haneda K, and Inazu T. Chemo-enzymatic synthesis of glycosylated insulin using a GlcNAc tag. Bioorg Med Chem. 2010;18(3):1259-1264.
Umekawa M, Li C, Higashiyama T, Huang W, Ashida H, Yamamoto K, Wang LX. Efficient glycosynthase mutant derived from Mucor hiemalis endo-β-N-acetylglucosaminidase capable of transferring oligosaccharide from both sugar oxazoline and natural N-glycan. J Biol Chem. 2010;285(1):511-521.
Ochiai H, Shimoda T, Fukae K, Maeda M, Ishii K, Yoshida K, Tezuka K, Tazuru K, Saijo H, Asai H, Kanatani A, Nishiuchi Y. Better peptides via chemical glycosylation: somatostatin analogues having a human complex-type N-glycan with improved drug properties. Chemistry. 2023;29(31):e202300111.
Huang W, Giddens J, Fan SQ, Toonstra C, Wang LX. Chemoenzymatic glycoengineering of intact IgG antibodies for gain of functions. J Am Chem Soc. 2012;134(29):12308-12318.
Iwamoto M, Sekiguchi Y, Nakamura K, Kawaguchi Y, Honda T, Hasegawa J. Generation of efficient mutants of endoglycosidase from Streptococcus pyogenes and their application in a novel one-pot transglycosylation reaction for antibody modification. PLoS One. 2018;13(2):e0193534.
Li T, Tong X, Yang Q, Giddens JP, Wang LX. Glycosynthase mutants of endoglycosidase S2 show potent transglycosylation activity and remarkably relaxed substrate specificity for antibody glycosylation remodeling. J Biol Chem. 2016;291(32):16508-16518.
Shivatare SS, Huang LY, Zeng Y , Liao JY, You T , Wang SY, Cheng T, Chiu CW, Chao P, Chen LT, Tsai TI, Huang CC, Wu CY, Lin NH, Wong C . Development of glycosynthases with broad glycan specificity for the efficient glyco-remodeling of antibodies. Chem Commun (Camb). 2018;54(48):6161-6164.
Hsieh YC, Guan HH, Lin CC, Huang TY, Chuankhayan P, Chen NC, Wang NH, Hu PL, Tsai YC, Huang YC, Yoshimura M, Lin PJ, Hsieh YH, Chen CJ. Structure-based high-efficiency homogeneous antibody platform by endoglycosidase Sz provides insights into its transglycosylation mechanism. JACS Au. 2024;4(6):2130-2150.
Parsons TB, Struwe WB, Gault J, Yamamoto K, Taylor TA, Raj R, Wals K, Mohammed S, Robinson CV, Benesch JL, Davis BG. Optimal synthetic glycosylation of a therapeutic antibody. Angew Chem Int Ed Engl. 2016;55(7):2361-2367.
Manabe S, Yamaguchi Y, Matsumoto K, Fuchigami H, Kawase T, Hirose K, Mitani A, Sumiyoshi W, Kinoshita T, Abe J, Yasunaga M, Matsumura Y, Ito Y. Characterization of antibody products obtained through enzymatic and nonenzymatic glycosylation reactions with a glycan oxazoline and preparation of a homogeneous antibody–drug conjugate via Fc N-glycan. Bioconjug Chem. 2019;30(5):1343-1355.
Ivanova A, Falcioni F. Challenges and opportunities for the large-scale chemoenzymatic glycoengineering of therapeutic N-glycosylated monoclonal antibodies. Front Catal. 2022;1:810779.
Li T, Li C, Quan DN, Bentley WE, Wang LX. Site-specific immobilization of endoglycosidases for streamlined chemoenzymatic glycan remodeling of antibodies. Carbohydr Res. 2018;458-459:77-84.
Chuang HY, Huang CC, Hung TC, Huang LY, Chiu CW, Chu KC, Liao JY, You TH, Wu CY, Chao P, Shivatare SS, Zeng YF, Tsai CS, Lin NH, Wu CY. Development of biotinylated and magnetic bead-immobilized enzymes for efficient glyco-engineering and isolation of antibodies. Bioorg Chem. 2021;112:104863.
Zhao K, Tang F, Shi W, Hong H, Zhou Z, Huang W, Wu Z. One-step immobilization and purification of genetic engineering CBD fusion EndoS on cellulose for antibodies Fc-glycan remodeling. Bioorg Chem. 2019;91:103114.
Muto H, Ishii N, Iwamoto M, Matsuo I. Rapid preparation of a glycan oxazoline and a homogeneously glycosylated antibody with an enzyme-immobilized monolithic column. Carbohydr Res. 2024;536:109024.
Zhang X, Ou C, Liu H, Wang LX. Synthesis and evaluation of three azide-modified disaccharide oxazolines as enzyme substrates for single-step Fc glycan-mediated antibody-drug conjugation. Bioconjug Chem. 2022;33(6):1179-1191.
Shi W, Li W, Zhang J, Li T, Song Y, Zeng Y, Dong Q, Lin Z, Gong L, Fan S, Tang F, Huang W. One-step synthesis of site-specific antibody-drug conjugates by reprograming IgG glycoengineering with LacNAc-based substrates. Acta Pharm Sin B. 2022;12(5):2417-2428.
Shoda S. Oxazoline donors enable effective glycoremodeling. Glycoforum. 2023;27(2): A6.
Higuchi Y, Eshima Y, Huang Y, Kinoshita T, Sumiyoshi W, Nakakita SI, Takegawa K. Highly efficient transglycosylation of sialo-complex-type oligosaccharide using Coprinopsis cinerea endoglycosidase and sugar oxazoline. Biotechnol Lett. 2017;39(1):157-162.
Eshima Y, Higuchi Y, Kinoshita T, Nakakita S, Takegawa K. Transglycosylation activity of glycosynthase mutants of endo-β-N-acetylglucosaminidase from Coprinopsis cinerea. PLoS One. 2015;10(7):e0132859.
Murakami S, Takaoka Y, Ashida H, Yamamoto K, Narimatsu H, Chiba Y. Identification and characterization of endo-β-N-acetylglucosaminidase from methylotrophic yeast Ogataea minuta. Glycobiology. 2013;23(6):736-744.
Nishizawa H, Iwamoto M, Ono Y. Identification and characterization of a novel thermo-stable endo-β-N-acetylglucosaminidase from Rhizomucor pusillus. J Biosci Bioeng. 2022;134(4):295-300.
Sjögren J, Struwe WB, Cosgrave EF, Rudd PM, Stervander M, Allhorn M, Hollands A, Nizet V, Collin M. EndoS2 is a unique and conserved enzyme of serotype M49 group A Streptococcus that hydrolyses N-linked glycans on IgG and α1-acid glycoprotein. Biochem J. 2013;455(1):107-118.
Shadnezhad A, Naegeli A, Sjögren J, Adamczyk B, Leo F, Allhorn M, Karlsson NG, Jensen A, Collin M. EndoSd: an IgG glycan hydrolyzing enzyme in Streptococcus dysgalactiae subspecies dysgalactiae. Future Microbiol. 2016;11:721-736.
Pier GB, Madin SH. Streptococcus iniae sp. nov., a beta-hemolytic Streptococcus isolated from an Amazon freshwater dolphin, Inia geoffrensis. Int J Syst Evol Microbiol. 1976;26(4):545-553.