Fairbanks AJ. The ENGases: versatile biocatalysts for the production of homogeneous N-linked glycopeptides and glycoproteins. Chem. Soc. Rev. 2017 Aug 14;46(16):5128-5146.
Kurogochi M. Glycan remodeling of glycoproteins using ENGases. Trends in Glycosci. Glycotech. 2018 Nov; 30(177):E209-E219.
Kurogochi M, Mori M, Osumi K, Tojino M, Sugawara S, Takashima S, Hirose Y, Tsukimura W, Mizuno M, Amano J, Matsuda A, Tomita M, Takayanagi A, Shoda S, Shirai T. Glycoengineered monoclonal antibodies with homogeneous glycan (M3, G0, G2, and A2) using a chemoenzymatic approach have different affinities for FcγRIIIa and variable antibody-dependent cellular cytotoxicity activities. PLoS One. 2015 Jul 22;10(7):e0132848.
Imai-Nishiya H, Mori K, Inoue M, Wakitani M, Iida S, Shitara K, Satoh M. Double knockdown of α1,6-fucosyltransferase (FUT8) and GDP-mannose 4,6-dehydratase (GMD) in antibody-producing cells: a new strategy for generating fully non-fucosylated therapeutic antibodies with enhanced ADCC. BMC Biotechnol. 2007 Nov 30;7:84.
Liu W, Padmashali R, Monzon OQ, Lundberg D, Jin S, Dwyer B, Lee YJ, Korde A, Park S, Pan C, Zhang B. Generation of FX-/- and Gmds-/- CHOZN host cell lines for the production of afucosylated therapeutic antibodies. Biotechnol. Prog. 2021 Jan;37(1):e3061.
Nason R, Büll C, Konstantinidi A, Sun L, Ye Z, Halim A, Du W, Sørensen DM, Durbesson F, Furukawa S, Mandel U, Joshi HJ, Dworkin LA, Hansen L, David L, Iverson TM, Bensing BA, Sullam PM, Varki A, de Vries E, de Haan CAM, Vincentelli R, Henrissat B, Vakhrushev SY, Clausen H, Narimatsu Y. Display of the human mucinome with defined O-glycans by gene engineered cells. Nat. Commun. 2021 Jul 1;12(1):4070.
Takashima S, Kurogochi M, Tsukimura W, Mori M, Osumi K, Sugawara SI, Amano J, Mizuno M, Takada Y, Matsuda A. Preparation and biological activities of anti-HER2 monoclonal antibodies with multibranched complex-type N-glycans. Glycobiology. 2021 Nov 18;31(10):1401-1414.
Henrissat B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991 Dec 1;280(Pt 2):309-316.
Drula E, Garron ML, Dogan S, Lombard V, Henrissat B, Terrapon N. The carbohydrate-active enzyme database: functions and literature. Nucleic Acids Res. 2022 Jan 7;50(D1):D571-D577.
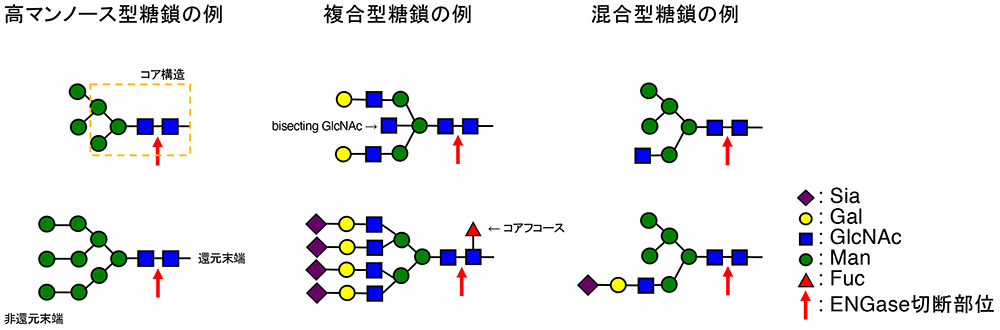
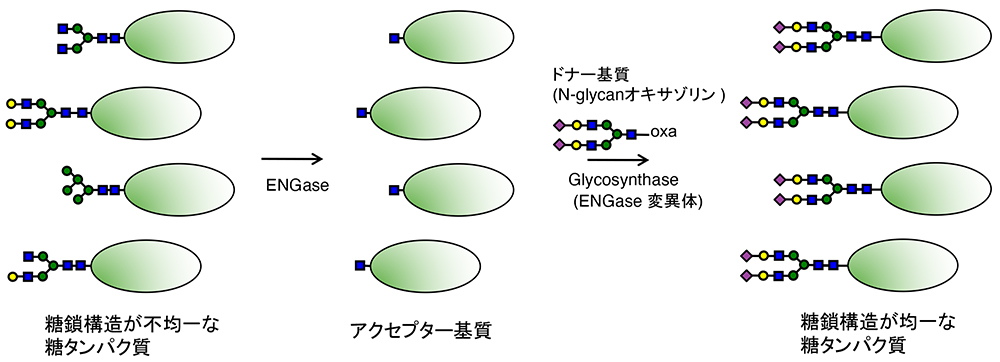
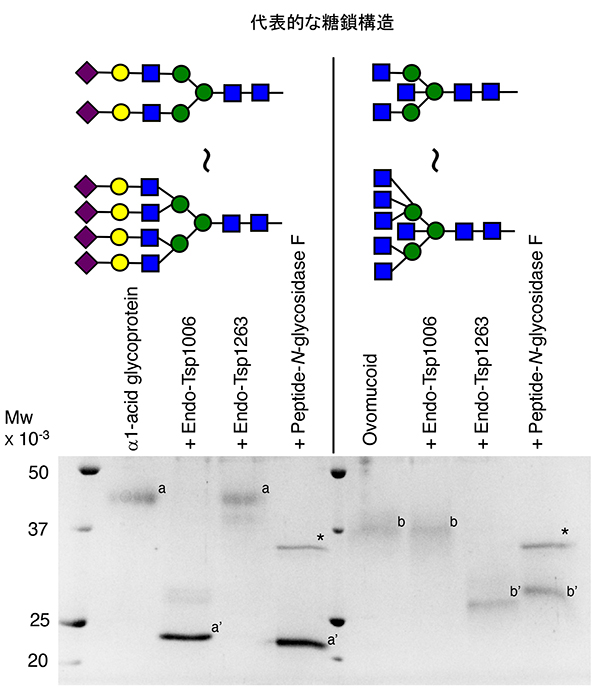
Yamamoto K, Chiba Y. Glycoengineering with ENGase. Glycoforum. 2026 Feb 2;29(1):A1.
Katoh T, Katayama T, Tomabechi Y, Nishikawa Y, Kumada J, Matsuzaki Y, Yamamoto K. Generation of a mutant Mucor hiemalis endoglycosidase that acts on core-fucosylated N-glycans. J. Biol. Chem. 2016 Oct 28;291(44):23305-23317.
Yamashita K, Kamerling JP, Kobata A. Structural study of the carbohydrate moiety of hen ovomucoid. Occurrence of a series of pentaantennary complex-type asparagine-linked sugar chains. J. Biol. Chem. 1982 Nov 10; 257(21):12809-12814.
Taguchi T, Seko A, Kitajima K, Muto Y, Inoue S, Khoo KH, Morris HR, Dell A, Inoue Y. Structural studies of a novel type of pentaantennary large glycan unit in the fertilization-associated carbohydrate-rich glycopeptide isolated from the fertilized eggs of Oryzias latipes. J. Biol. Chem. 1994 Mar 25; 269(12):8762-8771.
Liu CP, Tsai TI, Cheng T, Shivatare VS, Wu CY, Wu CY, Wong CH. Glycoengineering of antibody (Herceptin) through yeast expression and in vitro enzymatic glycosylation. Proc. Natl. Acad. Sci. USA. 2018 Jan 23;115(4):720-725.
Ito K, Okada Y, Ishida K, Minamiura N. Human salivary Endo-β-N-acetylglucosaminidase HS specific for complex type sugar chains of glycoproteins. J. Biol. Chem. 1993 Jul 25;268(21):16074-16081.
Miyagawa K, Matsumoto M, Yabuno S, Kawakami N, Hamaguchi T, Iizuka M, Minamiura N, Ito K. Transglycosylation of asparagine-linked complex-type oligosaccharides from glycoproteins by endo-β-N-acetylglucosaminidase HS. J. Appl. Glycosci. 2007;54(2):139-146.
Takashima S, Kurogochi M, Osumi K, Sugawara SI, Mizuno M, Takada Y, Amano J, Matsuda A. Novel endo-β-N-acetylglucosaminidases from Tannerella species hydrolyze multibranched complex-type N-glycans with different specificities. Glycobiology. 2020 Oct 21;30(11):923-934.
Bienes KM, Tautau FAP, Mitani A, Kinoshita T, Nakakita SI, Higuchi Y, Takegawa K. Characterization of novel endo-β-N-acetylglucosaminidase from Bacteroides nordii that hydrolyzes multi-branched complex type N-glycans. J. Biosci. Bioeng. 2022 Jul;134(1):7-13.
Doi K, Mitani A, Nakakita SI, Higuchi Y, Takegawa K. Characterization of novel endo-β-N-acetylglucosaminidases from intestinal Barnesiella intestinihominis that hydrolyze multi-branched complex type N-glycans. J. Biosci. Bioeng. 2024 Feb;137(2):101-107.